Appendix: Classification
Protophyta, Protozoa: One‑celled Plants and Animals
T HE SIMPLEST FORMS of cellular life are the one‑celled plants (Protophyta) and one‑celled animals (Protozoa). In both groups, however, there are many forms that defy attempts to place them definitely in one category or another because they display characteristics usually considered animal‑like along with others usually thought definitive of plants. The Dinoflagellata form such an indeterminate group, and are claimed both by zoologists and by botanists. Although a few are large enough to be seen without magnification, most are smaller. Some wear shells with spines and elaborate markings. Some have a remarkable, eye‑like sense organ. All dinoflagellates are immensely important in the economy of the sea as food for certain fishes and other animals. Noctiluca is a relatively large dinoflagellate of coastal waters, where it produces brilliant displays of phosphorescence, or by day reddens the water Sphaerella the abundance of its pigmented cells. Other species are the cause of the phenomenon known as “red tide,” in which the sea is discolored and fishes and other animals die from poisons given off by the minute cells. The red or green scum of high tide pools, “red rain,” and “red snow” are growths of these forms, or of green algae (e.g., Sphaerella). Much phosphorescence or “burning” of the sea is caused by dinoflagellates, which create a uniformly diffused light, lacking large spots of illumination. Examined closely, in a vessel of water, the light is seen to consist of tiny sparks.


The Radiolaria are one‑celled animals whose protoplasm is contained in siliceous shells of extraordinary beauty. These minute shells, sinking to the bottom, accumulate there to form one of the characteristic oozes or sediments of the sea floor. The Foraminifera are another unicellular group. Most have calcareous shells, though some build their protective structures with sand grains or sponge spicules. The shells, eventually drifting to the floor of the ocean, cover vast areas with calcareous sediments that, through geologic change, may become compacted into limestone or chalk, and raised to form such features of the present landscape as the chalk cliffs of England. Most Foraminifera are so minute that one gram of sand might contain up to 50,000 shells. On the other hand, a fossil species, Nummulites, was sometimes 6 or 7 inches across and formed limestone beds in Northern Africa, Europe, and Asia. This limestone was used in the building of the Sphinx and the great pyramids. Fossil Foraminifera are much used by geologists in the oil industry in correlating rock strata.
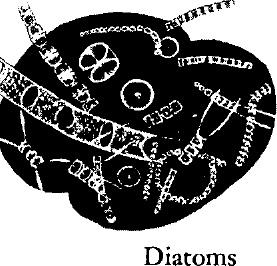
Diatoms (Greek, diatomos –cut in two) are minute plants usually classified among the yellow‑green algae because they contain granules of yellow pigment. They exist as single cells or in chains of cells. The living tissue of a diatom is encased within a shell of silica, of which one half fits over the other, as a lid over a box. Fine etchings on the surface of the shell create beautiful patterns and are characteristic for the various species. Most diatoms live in the open sea, and because they exist in inconceivable abundance are the most important single food stuff in the ocean, being eaten not only by many small animals of the plankton, but by many larger creatures, as mussels and oysters. The hard shells sink to the bottom after the death of the tissues, and accumulate there to form diatom oozes that cover vast areas of ocean floor.

The blue‑green algae, or Cyanophyceae, are among the simplest and oldest forms of life and are the most ancient plants that still exist. They are widely distributed and occur even in hot springs and other places where conditions are so difficult that no other plant life can exist. They often multiply in phenomenal numbers, giving the surface of ponds and other still waters a colored film known as water bloom. Most are encased in gelatinous sheaths that protect them from extreme heat or cold. They are well represented in the “black zone” above high‑tide line on rocky shores.
Thallophyta: Higher Algae
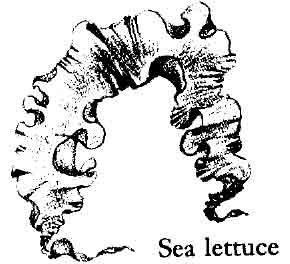
T HE GREEN ALGAE, or Chlorophyceae, are able to endure strong light and thrive high in the intertidal zone. They include such familiar forms as the leafy sea lettuce and a stringy, tube‑like alga of high rocks and tide pools called Enteromorpha (“intestine‑shaped”). In the tropics some of the most common green algae are the brush‑shaped Penicillus that forms minute groves over the coral reef flats, and the beautiful little cup alga, Acetabularia, like tiny, inverted mushrooms of purest green. Some of the green algae of the tropics are important in the economy of the sea as concentrators of calcium. Although the group is most typical of warm, tropical seas, the green algae are found on the shore wherever there is strong sunlight, and others of the group live in fresh water.
The brown algae, or Phaeophyceae, possess various pigments that conceal their chlorophyll, so their prevailing colors are brown, yellowish, or olive‑green. They are largely absent from warmer latitudes except in deep water, being unable to endure heat and strong sun. An exception is the Sargassum weed of tropical shores, which drifts northward in the Gulf Stream. On northern coasts the brown rockweeds live between tide lines, and the kelps or oarweeds from the low‑tide line down to depths of 40 to 50 feet. Although all of the algae select and concentrate in their tissues many different chemicals present in sea water, the brown seaweeds and especially the kelps are extraordinary in the quantity of iodine stored. Formerly they were utilized widely in the industrial production of iodine. The same seaweeds now are important in the production of the carbohydrate algin for use in fire‑resistant textiles, jellies, ice cream, cosmetics, and various industrial processes. The presence of alginic acid gives these seaweeds their great resilience in heavy surf.

The red algae, or Rhodophyceae, most sensitive light of all the seaweeds, send only a few hardy species (including Irish moss and dulse) into the intertidal zone; most are delicate and graceful seaweeds living for the most part below low water. Some live deeper than any other seaweeds, going down into the dim regions 200 fathoms or more below the surface. Some (the corallines) form hard crusts on rocks or shells. Containing magnesium carbonate as well as calcium carbonate, these algae seem to have played an important geochemical role in earth history, perhaps having aided the formation of the magnesium‑rich marble dolomite.

Porifera: Sponges
T HE SPONGES (Porifera, or pore‑bearers) are among the simplest of animals, being little more than an aggregation of cells. Yet they have gone a step beyond the Protozoa, for there are inner and outer layers of cells, with some hint of specialization of function–some for drawing in water, some for taking in food, some for reproduction. All these cells cohere and work together to carry out the single purpose of the sponge–to pass the waters of the sea through the sieves of its own being. A sponge is an elaborate system of canals contained in a matrix of fibrous or mineral substance, the whole pierced by numerous small entrance pores and larger exit holes. The inmost or central cavities are lined with flagellated cells that remind one of protozoan flagellates. The lashing of the whiplike flagella creates currents to draw in water. In passage through the sponge, the water gives up food, minerals, and oxygen, and carries away waste products.

To a certain extent, each of the smaller groups within the sponge phylum has a physical appearance and habit of life that is characteristic, yet the sponges are probably more plastic in relation to their environment than any other animals. In surf they take the form of a flattened crust, almost without regard to species; in deep, quiet water they may assume an upright tubular form, or branch in a way suggestive of shrubbery. Their shape, therefore, is little or no aid in identification, and the classification of sponges is based chiefly on the nature of their skeleton, which is a loose network of minute hard structures called spicules. In some the spicules are calcareous. In others they are siliceous, although sea water contains only a trace of silica and the sponge must have to filter prodigious quantities to obtain enough for its spicules. The function of extracting silica from sea water is confined to primitive forms of life, and among animals does not occur above the sponges. Commercial sponges fall into a third group, having a skeleton of horny fibers. They are confined to tropical waters.

From such a beginning toward specialization, nature seems to have gone back and made a fresh start with, other materials. All evidence points toward a separate origin for the coelenterates and all other more complex animals, leaving the sponges in an evolutionary blind alley.
Coelenterata: Anemones, Corals, Jellyfish, Hydroids
T HE COELENTERATES, despite their simplicity, foreshadow the basic plan on which, with elaborations, all the more highly developed animals are formed. They possess two distinct layers of cells, the outer ectoderm and the inner endoderm, sometimes with an undifferentiated middle layer that is not cellular but is the forerunner of the third cell layer, the mesoderm, of the higher groups. Each coelenterate is basically a hollow double‑walled tube, closed at one end and open at the other. Variations of this plan have resulted in such diverse forms as the sea anemones, corals, jellyfish, and hydroids.
All coelenterates possess stinging cells called nematocysts, each of which is a coiled, pointed thread contained in a sac of turgid fluid, ready to be expelled to impale or entangle passing prey. Stinging cells are not developed in higher animals; although they have been reported in flatworms and sea slugs, they have been secondarily acquired by eating coelenterates.

The Hydrozoa display most clearly another peculiarity of this group, known as alternation of generations. An attached, plantlike generation produces a medusoid generation, shaped like small jellyfish. These, in turn, produce another plantlike generation. In the hydroids the more conspicuous generation is an attached, branching colony bearing tentacled individuals, or hydranths, on its “stems.” Most of these are shaped like small sea anemones and capture food. Other individuals bud off the new generation–tiny medusae that (in many forms) swim away, mature, and shed eggs or sperm cells into the sea. An egg produced by such a medusa, when fertilized, develops into another plantlike stage.

In another group, the Scyphozoa, or true jellyfish, the plantlike generation is the inconspicuous one, and the medusae are highly developed. The jellyfish range from very small creatures to the immense arctic jelly, Cyanea, which reaches an extreme diameter of 8 feet (1 to 3 feet is more common) with tentacles up to 75 feet long.
In the Anthozoa (flower animals) the medusoid generation has been completely lost. This group includes the anemones, corals, sea fans, and sea whips. The anemone represents the basic plan; all the rest of this group are colonial forms in which the individual, anemone‑like polyps are embedded in some sort of matrix, which may be stony, as in the reef‑building corals, or, in the sea fans and sea whips, may consist of a horny substance of protein nature, similar to the keratin of vertebrate hair, nails, and scales.

Ctenophora: Comb Jellies
THE ENGLISH WRITER Barbellion once said that a comb jelly in sunlight is the most beautiful thing in the world. Its tissues are almost crystal clear, and as this little ovoid creature twirls in the water it flashes iridescent lights. The ctenophores, or comb jellies, are sometimes mistaken for jellyfish because of their transparency, but there are various structural differences, with the “comb‑plates” being characteristic of the phylum. These occur in eight rows on the outer surface. Each plate has a hinged attachment and bears hairlike cilia along its free edge; as the plates flash in succession to propel the animal through the water, the cilia break up the rays of sunlight and produce the characteristic flashing.

Like some of the jellyfish, most ctenophores possess long tentacles. These are equipped not with stinging cells, but with sticky pads that capture prey by entanglement. Ctenophores eat enormous numbers of fish fry and other small animals. They live chiefly in the surface waters.
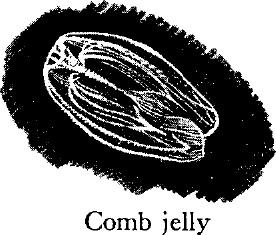
The ctenophores comprise a small phylum, with less than 100 species. Members of one of their groups have flattened bodies and do not swim, but creep on the ocean floor. Some specialists believe these creeping ctenophores have given rise to the flatworms.
Platyhelminthes: Flatworms
THE FLATWORMS include many parasitic as well as many free‑living forms. Leafy thin, the free‑living flatworms flow like a living film over rocks or sometimes swim by flapping undulations in a way reminiscent of skates. They have made significant advances in an evolutionary sense. They are the first to possess three primary layers of cells, a characteristic of all higher animals. They also have a bilateral type of symmetry (one side being a mirror image of the other), with a head end that always goes first. They have the simple beginnings of a nervous system and eyes that may be only simple pigment spots or, in some species, well‑developed organs with lenses. There is no circulatory system, and perhaps it is because of this that all flatworms have such thin bodies, in which all parts are in easy communication with the exterior, and oxygen and carbon dioxide are easily passed through surface membranes to underlying tissues.

Flatworms are found among seaweeds, on rocks, in tide pools, and lurking in dead mollusk shells. They are usually carnivorous, devouring worms, crustaceans, and mollusks of minute size.
Nemertea: Ribbon Worms
THE RIBBON WORMS have extraordinarily elastic bodies, sometimes round, sometimes flat. One of them, the bootlace worm (Lineus longissimus ) of British waters, may attain a length of 90 feet and is the longest of all the invertebrates. The American Cerebratulus of shallow coastal waters often is 20 feet long and about an inch wide. Most, however, are only a few inches long and many are considerably less than an inch. They habitually contract into coils or knots when disturbed.
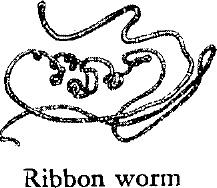
All ribbon worms are highly muscular but lack the co‑ordination of nerve and muscle that higher worms have. There is a brain consisting of simple nerve ganglia. Some have primitive hearing organs, and the characteristic slits along the sides of the head (suggestive of a mouth) seem to contain important organs of sensation. Although there are a few hermaphroditic species, in most ribbon worms the sexes are separate. There is, however, a strong tendency toward asexual reproduction, and associated with this is a habit of breaking up into many pieces when handled. The fragments then regenerate complete worms. Professor Wesley Coe of Yale University found that a certain species of ribbon worm could be cut repeatedly until eventually miniature worms less than one one‑hundred thousandth the volume of the original were obtained. An adult can live a year without food, according to Professor Coe, compensating for lack of nourishment by diminishing in size.
The ribbon worms are unique in the possession of an extensible weapon called a proboscis, enclosed in a sheath and capable of being suddenly everted, hurled out, and coiled around the prey, which is then drawn back toward the mouth. In many species the proboscis is armed with a sharp lance, or stylet, which if lost is quickly replaced by another held in reserve. All ribbon worms are carnivorous, and many prey on the bristle worms.
Annelida: Bristle Worms
THE ANNELID (ringed, or segmented) worms include several classes, one of which, the Polychaeta (many bristles) includes most marine annelids. Many of the polychaetes, or bristle worms, are active swimmers that make their living as predators; others are more or less sedentary, building tubes of various sorts in which they live, either feeding on detritus in sand or mud or on plankton which they strain from the water. Some of these worms are among the most beautiful creatures of the sea, their bodies shining with iridescent splendor, or adorned with feathery crowns of tentacles in soft and beautiful colors.

In their structure they represent a great advance over lower forms. Most of them possess a circulatory system (although the blood worm, Glycera, much used as bait, has no blood vessels but a blood‑filled cavity between the skin and the alimentary canal) and so are able to dispense with the thinness of body of the flatworms, for the blood flowing through vessels transports food and oxygen to all parts of the body. The blood is red in some, green in others. The body consists of a series of segments, several of the anterior ones being fused to form the head. Each segment bears a pair of unbranched, unsegmented paddle‑like appendages for crawling or swimming.
Bristle worms include many diverse forms. The familiar nereids, or clam worms, often used for bait, spend most of their lives in crude burrows among stones on the sea bottom but emerge to hunt or, in swarms, to spawn. The sluggish scale worms live under rocks, in muddy burrows, or among the holdfasts of seaweeds. The serpulid worms build variously shaped limy tubes from which only their heads emerge; other worms, like the beautifully plumed Amphitrite, form mucous tubes under rocks or crusts of coralline algae or on muddy bottoms, and a worm of colonial habit, Sabellaria, uses coarse sand grains to build elaborate structures that may be several feet across. Though honeycombed with the burrows of the worms, these massive dwelling places are strong enough to bear the weight of a man.
Arthropoda: Lobsters, Barnacles, Amphipods
THE ARTHROPOD (jointed foot) phylum is an enormous group, comprising five times as many species as are included in all the rest of the animal phyla combined. The arthropods include the crustacea (e.g., crabs, shrimps, lobsters), the insects, the myriopods (centipedes and millipedes), the arachnids (spiders, mites, and king crabs) and the tropical, wormlike Ony‑chophora. All marine arthropods belong to the class Copepod Crustacea except for a scant handful of insects, a few mites and sea spiders, and the king crabs.

Whereas the paired appendages of the annelids are simple flaps, those of the arthropods possess multiple joints and are specialized to perform such varied functions as swimming, walking, handling food, and gaining sensory impressions of the environment. Whereas the annelids interpose only a simple cuticle between their internal organs and the environment, the arthropods protect themselves by a rigid skeleton of chitin impregnated with lime salts. This, in addition to being protective, has the advantage of giving a firm support for the insertion of muscles. On the other hand there is the disadvantage that, as the animal grows, the rigid outer covering must be shed from time to time.
The crustaceans include such familiar animals as crabs, lobsters, shrimps, and barnacles, as well as less‑known creatures like the ostracods, isopods, amphipods, and copepods, all of which are important or interesting for one reason or another.
The ostracods are unusual arthropods in that they are not segmented but are enclosed in a two‑part carapace, or shell, flattened from side to side, and opened and closed by muscles like a mollusk’s shell. The antennae act as oars and are extended through the opened carapace to row the little animal through the water. Ostracods often live in seaweeds or in sand on the ocean floor, usually being quiet by day and coming out to feed at night. Many marine ostracods are luminous and as they swim about emit little puffs of bluish light. They are one of the chief sources of phosphorescence at sea. Even when dead and dried they retain the phosphorescent quality to an astonishing degree. Professor E. Newton Harvey of Princeton University says in his authoritative volume Bioluminescence that during the Second World War Japanese army officers used dried ostracod powder in advanced positions where use of flashlights was prohibited–by adding a few drops of water to a little powder in the palm of the hand, they could obtain enough light to read dispatches.

Copepods (oar‑footed) are very small crustaceans with rounded bodies, jointed tails, and oarlike legs with which to propel themselves jerkily. In spite of their minute size (from microscopic to half‑inch) the copepods form one of the basic populations of the sea, and are food for an immense variety of other animals. They are an indispensable link in the food chain by which the nutrient salts of the sea are eventually made available (via plant plankton, animal plankton, carnivores) to larger animals such as fishes and whales. Copepods of the genus Calanus, known as “red feed,” redden large areas of ocean surface and are eaten in prodigious numbers by herring and mackerel and also by certain whales. Birds of the open sea such as petrels and albatrosses are plankton feeders and sometimes subsist largely on copepods. In their turn, the copepods graze on diatoms, eating sometimes as much as their own weight in a day.
Amphipods are small crustaceans that are flattened from side to side, while isopods are flattened from upper to lower surface. The names are a scientific reference to the kinds of appendages possessed by these small creatures. The amphipods have feet that can be used both for swimming and walking or crawling. The isopods, or “equal‑footed” animals, have appendages that show little difference in size and shape from one end of the body to the other.
On the shore the amphipods include the beach hoppers, or sand fleas, that rise in clouds (leaping, not flying) from masses of seaweed when they are disturbed, and others that live offshore in seaweed and under rocks. They eat fragments and bits of organic debris and are themselves eaten in great number by fish, birds, and other larger creatures. Many amphipods wriggle along on their sides when out of water. Sand hoppers use their tails and posterior legs as a spring and progress by leaps; other species swim.

Isopods of the shore (closely related to the familiar sow bugs of the garden) include the slaters (sea roaches, wharf rats, quay lice) often seen running over rocks and wharf pilings. These have left the water and seldom return to it; they drown if long submerged. Others live offshore, often in seaweeds whose color and form they mimic. Still others swarm in tide pools, sometimes nipping the skin of human waders to produce a tingling or itching sensation. Most are scavengers; some are parasites; and some form habitual associations (commensalism) with an animal of unrelated species.

Both amphipods and isopods carry their young in brood chambers instead of liberating eggs into the sea. This habit has helped some in each group to live high on the shore and is a necessary preliminary to land existence.
The barnacles belong to the order Cirripedia (Latin, cirrus –a ringlet or curl), presumably named because of their gracefully curving feathery appendages. The larval stages are free‑living and resemble the larvae of many other crustaceans, but the adults are attached, living in a shell of calcareous material, fixed to rocks or other hard objects. The gooseneck barnacles are attached by a leathery stalk; the rock or acorn barnacles are attached directly. The gooseneck barnacles are often oceanic, attaching themselves to ships and floating objects of all sorts. Some of the acorn barnacles grow‑on the hide of whales or the shells of sea turtles.
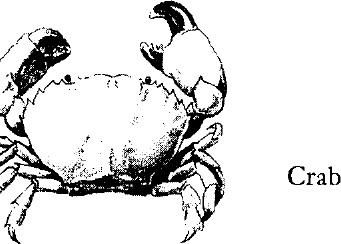
The large crustaceans–shrimps, crabs, and lobsters–not only are most familiar but display the typical arthropod plan of body most clearly. The head and thoracic regions usually are fused and covered with a hard shell, or carapace; only the appendages indicate the division into segments. The flexible abdomen or “tail,” on the other hand, is divided into segments and usually is an important aid to swimming. Crabs, however, keep the tail segments folded under the body.
The hard shell of an arthropod must be shed periodically as the animal grows. The creature gets out of the old shell through a slit that opens up usually across the back. Underneath is the new shell, much folded and wrinkled, soft and tender. The crustacean, after shedding, may spend days in seclusion, hiding from enemies until its armor has hardened.
The class Arachnoidea includes in one group the horseshoe crabs, and in another diverse one the spiders and mites, only a few of which are marine. The horseshoe, or king, crab has a peculiar distribution, being very abundant on the Atlantic coast of America, absent from Europe, and represented by three species on the Asiatic coast from India to Japan. Its larval stages closely resemble the ancient trilobites of Cambrian times and as a reminder of those past ages it is often called a living fossil. Horseshoe crabs are abundant along the shores of bays and other relatively quiet waters, where they eat clams, worms, and other small animals. They come out on beaches early in the summer to lay eggs in depressions scooped out in the sand.

Bryozoa: Moss Animals, Sea Laces
THE BRYOZOA are a group of uncertain position and relationships, including rather diverse forms. They may appear as fluffy plantlike growths often mistaken for seaweeds, especially when found dried on the shore. Another form grows as flat hard patches encrusting seaweeds or rocks and having a lacy appearance. Still another type is a branched and upright growth of gelatinous texture. All of these are colonial forms or associations of many individual polyps, all living in adjoining cells or embedded in a unifying matrix.
The encrusting Bryozoa, or sea laces, are beautiful mosaics of closely set compartments, each inhabited by a small tentacled creature that superficially resembles the hydroid polyp, but possesses a complete digestive system, a body cavity, simple nervous system, and many other features of higher animals. The individuals of a bryozoan colony are largely independent of each other, instead of being connected as the hydroids are.

The Bryozoa are an ancient group dating from the Cambrian. They were considered seaweeds by early zoologists, and later were classified as hydroids. There are about 3000 marine species, compared with only about 35 in fresh waters.
Echinodermata: Starfish, Sea Urchins, Brittle Stars, Sea Cucumbers
OF ALL the invertebrates, the echinoderms are most truly marine, for among their nearly 5000 species not one lives in fresh water or on land. They are an ancient group, dating from the Cambrian, but in all the hundreds of millions of years since then none has even attempted to make the transition to a land existence.
The earliest echinoderms were the crinoids, or sea lilies, stalked forms that lived attached to the floor of Paleozoic seas. Some 2100 fossil species of crinoids are known, in contrast to about 800 living species. Today most crinoids live in East Indian waters; a few occur in the West Indian region and come as far north as Cape Hatteras, but there are none in the shallow waters of New England.
The common echinoderms of the shore represent the four remaining classes of the phylum: the sea stars, the brittle and serpent stars, the sea urchins and sand dollars, and the holothurians, or sea cucumbers. In all members of the group there is a recurrent insistence on the number five, many of the structures occurring in fives or multiples of five, so that the figure is almost a symbol of the group.
The sea stars, or starfish, have flattened bodies, many in the conventional five‑pointed shape, though the number of arms varies. The skin is roughened by hard limy plates from which short spines grow. In most species the skin also bears structures like minute forceps on flexible stalks (called pedicellaria); with these the animal keeps the skin clear of sand grains and also picks off larvae of sedentary forms that try to settle there. This is necessary because the delicate breathing organs–soft rosettes of tissue–also project through the skin.

Like all other echinoderms, the starfish possess a so‑called water‑vascular system that functions in locomotion and secondarily in other ways, and consists of a series of water‑filled tubes running to all parts of the body. Intake of sea water is accomplished, in starfish, through a conspicuous perforated plate on the upper surface–the madreporite (mother of pores). The fluid passes along the water canals and eventually into the many short flexible tubes (tube feet) that occupy the long grooves on the under surface of the arms. Each tube bears a sucker at its tip. The tube feet can be lengthened or contracted by changes of hydrostatic pressure–when extended, the suckers grip the underlying rock or other hard surface and the animal pulls itself along. The tube feet are used also to grip the shells of mussels or other bivalve mollusks on which the starfish preys. As the starfish moves, any of its various arms may Brittle star go first and thus serve as temporary “head.”

In the slender, graceful brittle stars and serpent stars the arms are not grooved and the tube feet are reduced. However, these animals progress rapidly by writhing motions of the arms. They are active predators and feed on a variety of small animals. Sometimes they lie in “beds” of many hundreds of animals on the sea bottom offshore–a living net through which scarcely any small creature can safely reach bottom.
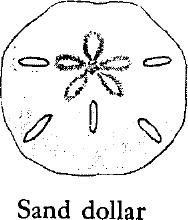
In the sea urchins the tube feet are arranged in five avenues or rows running from upper to lower apexes of the body, just as the meridians on a globe run from pole to pole. The skeletal plates of the urchins are articulated rigidly to form a globular shell, or test. The only movable structures are the tube feet, which are thrust out through perforations in the test, the pedicellaria, and the spines, which are mounted on protuberances on the plates. The tube feet are retracted when the animal is out of water, but when submerged they may be extended beyond the spines to grasp the substratum or to capture prey. They may also perform some sensory functions. In the various species the spines differ greatly in length and thickness.

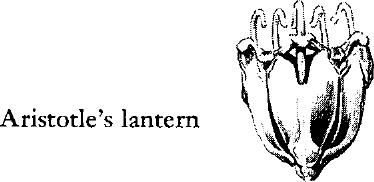
The mouth is on the under surface, surrounded by five white, shining teeth used to scrape vegetation off the rocks and also to assist in locomotion. (Although other invertebrates–e.g., the annelids–have biting jaws, the urchins are the first to have grinding or chewing organs.) The teeth are operated by an internally projecting apparatus of calcareous rods and muscles known to zoologists as Aristotle’s lantern. On the upper surface the digestive tract opens to the exterior through a centrally placed anal pore. Around this are five petal‑shaped plates, each bearing a pore that serves to discharge eggs or sperm. The reproductive organs are arranged in five clusters just under the upper or dorsal surface. They are practically the only soft parts the animal possesses and it is for these that the sea urchins are sought as human food, especially in Mediterranean countries. Gulls hunt the urchins for a similar purpose, often dropping them on the rocks to break the tests so that they can eat out the soft parts.

The eggs of the sea urchins have been used extensively in biological studies of the nature of the cell, and Jacques Loeb in 1899 used them in a historic demonstration of artificial parthenogenesis, causing an unfertilized egg to develop merely by treating it with chemicals or by mechanical stimulus.
The holothurians, or sea cucumbers, are curious echinoderms with soft, elongated bodies. They crawl on one surface with the mouth end foremost and so have secondarily substituted a functional bilateral symmetry for the radial symmetry characteristic of the phylum. Tube feet, where present, are confined to three rows on the functional under surface of the body. Some holothurians are burrowing forms, using small spicules embedded in the body surface to grasp the surrounding mud or sand and aid their progress. The shapes of these spicules vary with the species and often must be studied microscopically before correct identification can be made. The holothurians are large and abundant in tropical seas (they are the trepang, or beche‑de‑mer, of commerce) and in northern waters are represented by smaller species living on offshore bottoms or among intertidal rocks and seaweeds.
Mollusca: Clams, Snails, Squids, Chitons
BECAUSE OF their endlessly varied shells, often intricately made and beautifully adorned, some of the mollusks probably are better known than any other animals of the shore. As a group they possess qualities different from those of any other invertebrates, although their more primitive members and the nature of their larvae suggest that their remote ancestors may have resembled those of the flatworms. They have soft, unsegmented bodies typically protected by a hard shell. One of the most remarkable and characteristic molluscan structures is the mantle, a cloaklike tissue that encloses the body, secretes the shell, and is responsible for its complex structure and adornment.
The most familiar mollusks are the snail‑like gastropods and the clamlike bivalves. The most primitive mollusks are the creeping, sluggish coat‑of‑mail shells, or chitons, the least known are the tusk shells, or scaphopods, and the most highly developed class the cephalopods, represented by the squids.

The shells of the gastropods are univalve or in one piece, and coiled in more or less spiral fashion. Nearly all snails are “right‑handed,” that is, the opening is to the right as it faces the observer. One of the exceptions is the “left‑handed conch,” one of the most common gastropods of Florida beaches. Occasionally a left‑handed individual occurs in a normally right‑handed species. Some gastropods have reduced the shell to an internal remnant, as in the sea hares, or have lost it entirely, as in the sea slugs or nudibranchs (in which, however, a coiled shell is present in the embryo).
The snails are for the most part active animals, both the vegetarians that move about scraping plant food from the rocks and the carnivores that capture and devour animal prey. The sedentary boat shells, or slipper shells, are exceptions; they attach themselves to shells or to the sea bottom and live on diatoms strained from the water, in the manner of oysters, clams, and other bivalves. Most snails glide about on a flattened muscular “foot,” or they may use this same organ to burrow into the sand. When disturbed, or at low tide, they draw back into their shells, the opening being closed by a calcareous or horny plate called the operculum. The shape and structure of the operculum vary greatly in the different species and sometimes it is useful in identification. In common with other mollusks (except the bivalves) the gastropods have a remarkable, tooth‑studded band, the radula, on the floor of the pharynx, or, in some species, on the end of a long proboscis. The radula is used to scrape off vegetation or to drill holes in shelled prey.
The bivalves, with few exceptions, are sedentary. Some (e.g., the oyster) fix themselves permanently to a hard surface. Mussels and some others anchor themselves by secreting silklike byssus threads. The scallops and the lima clams are examples of the few bivalves that possess the ability to swim. The razor clams have a slender pointed foot by means of which they dig deeply and with incredible speed into the sand or mud.

Bivalves that bury deeply in the substratum are able to do so because they possess a long breathing tube, or siphon, through which they draw in water and so receive oxygen and food. Although most are suspension feeders, filtering minute food organisms from the water, some, including the tellins and coquina clams, live on detritus that accumulates on the sea floor. There are no carnivorous bivalves.
The shells of gastropods and bivalves are secreted by the mantle. The basic chemical material of molluscan shells is calcium carbonate, which forms the outer layer of calcite, and the inner layer of aragonite, which is a heavier and harder substance although it has the same chemical composition. Calcium phosphate and magnesium carbonate also are contained in mollusk shells. The limy materials are laid down on an organic matrix of conchiolin, a substance chemically allied to chitin. The mantle contains pigment‑forming cells as well as shell‑secreting cells. The rhythm of activity of these two kinds of cells results in the marvelous sculpturing and color patterns of molluscan shells. Although shell formation is affected by many factors in the environment and in the physiology of the animal itself, the basic hereditary pattern is so strongly determined that each species of mollusk has its characteristic shell by which it may be identified.
A third class of the mollusk phylum consists of the cephalopods, so unlike the snails and clams that superficially it is hard to reconcile the relationship. Although ancient seas were dominated by shelled cephalopods, all but one (the chambered nautilus) have now lost the external shell, retaining only an inconspicuous internal remnant. One large group, the decapods, have cylindrical bodies with ten arms; they are represented by the squids, the ramshorn shell, and the cuttlefish. Another group, the octopods, have baglike bodies with eight arms; examples are the octopus and the argonaut.
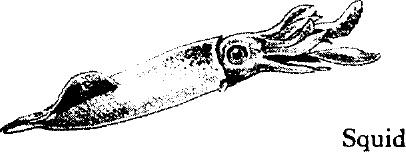
The squids are strong and agile; over short distances they are probably the swiftest animals of the sea. They swim by expelling a jet of water through the siphon, controlling the direction of motion by pointing the siphon forward or backward. Some of the smaller species swim in schools. All squids are carnivorous, preying on fish, crustaceans, and various small invertebrates. They are sought by cod, mackerel, and other large fish, and are a favorite bait. The giant squid is the largest of all invertebrates. The record specimen, taken on the Grand Banks of Newfoundland, measured about 55 feet including the arms.

Octopuses are nocturnal animals and, according to those most familiar with their habits, are timid and retiring. They live in holes or among rocks, feeding on crabs, mollusks, and small fish. Sometimes the location of an octopus den may be discovered by the pile of empty mollusk shells near the entrance.
The chitons belong to a primitive order of mollusks, the Amphineura. Most of them wear a shell consisting of eight transverse plates bounded by a tough band, or girdle. They creep sluggishly over rocks, scraping off vegetation. At rest, they settle into a depression, blending so well with their surroundings that they are easily overlooked. They are sought as food (sea beef) by West Indian natives.
The fifth class of mollusks consists of the little‑known scaphopods (tooth shells or tusk shells), which form shells resembling an elephant’s tusk, from one to several inches long and open at both ends. They dig into sandy bottoms, using a small, pointed foot. Some specialists think their structure may be similar to that of the ancestors of all mollusks. However, this is a field for speculation, since the principal classes of mollusks were all defined early in the Cambrian, and clues to the nature of the ancestral forms are exceedingly vague. The tooth shells number about 200 species, and are widely distributed in all seas. None, however, are intertidal.
Chordata: Subphylum Tunicata
THE ASCIDIANS, or sea squirts, are the most common representatives on the shore of that interesting group of early chordates, the Tunicata. As forerunners of the vertebrates, or backboned animals, all of the chordates have at some time a stiffening rod of cartilaginous material, an evolutionary forecast of the vertebral column which all the higher animals were to possess. The adult ascidian paradoxically suggests a creature of low and simple organization, with a physiology somewhat like that of oysters or clams. It is only in the larva that the chordate characteristics are clear. Though minute, the larva strongly resembles the tadpole of a frog, possessing a notochord and a tail and swimming actively. At the end of the larval period it settles down, becomes attached, and undergoes metamorphosis to the much simpler adult form, in which the chordate characters are lost. This is a curious phenomenon of evolution, which seems to be degenerative rather than progressive, with the larva displaying more advanced characteristics than the adult.

The adult sea squirt is shaped like a bag with two tubular openings or siphons for water intake and outgo, and a pharynx perforated with many slits through which water is strained. The common name refers to the fact that when the animal is disturbed it contracts sharply, forcing jets of water out through the siphons. In the so‑called simple ascidians the animals live as separate individuals, each enclosed in a tough covering or test of material chemically akin to cellulose. Sand and debris often adhere to this test, forming a mat in which the actual shape of the animal is seldom apparent. In this form they often grow profusely on wharf pilings, floats, and rocky ledges. In the compound, or colonial, type of ascidian many individuals live together, embedded in a tough gelatinous substance. Unlike a group of simple ascidians, the various individuals of a colony are derived by asexual budding from one individual, the founder of the colony. One of the commonest compound sea squirts is the sea pork, Amaroucium, named from the usually gray, gristly appearance of its colonies. These may form a thin mat on the under side of a rock or, offshore, grow erect, forming thick slabs that may break off and be carried in to shore. The individuals composing the colony are not easily seen, but under a lens pits in the surface appear, each the opening through which a single sea squirt communicates with the outside world. In the beautiful compound sea squirt Botryllus, however, the individuals form flowerlike clusters, easily visible.
Дата добавления: 2015-05-13; просмотров: 1979;