V. The Coral Coast
I DOUBT that anyone can travel the length of the Florida Keys without having communicated to his mind a sense of the uniqueness of this land of sky and water and scattered mangrove‑covered islands. The atmosphere of the Keys is strongly and peculiarly their own. It may be that here, more than in most places, remembrance of the past and intimations of the future are linked with present reality. In bare and jaggedly corroded rock, sculptured with the patterns of the corals, there is the desolation of a dead past. In the multicolored sea gardens seen from a boat as one drifts above them, there is a tropical lushness and mystery, a throbbing sense of the pressure of life; in coral reef and mangrove swamp there are the dimly seen foreshadowings of the future.
This world of the Keys has no counterpart elsewhere in the United States, and indeed few coasts of the earth are like it. Offshore, living coral reefs fringe the island chain, while some of the Keys themselves are the dead remnants of an old reef whose builders lived and flourished in a warm sea perhaps a thousand years ago. This is a coast not formed of lifeless rock or sand, but created by the activities of living things which, though having bodies formed of protoplasm even as our own, are able to turn the substance of the sea into rock.
The living coral coasts of the world are confined to waters in which the temperature seldom falls below 70° F. (and never for prolonged periods), for the massive structures of the reefs can be built only where the coral animals are bathed by waters warm enough to favor the secretion of their calcareous skeletons. Reefs and all the associated structures of a coral coast are therefore restricted to the area bounded by the Tropics of Cancer and Capricorn. Moreover, they occur only on the eastern shores of continents, where currents of tropical water are carried toward the poles in a pattern determined by the earth’s rotation and the direction of the winds. Western shores are inhospitable to corals because they are the site of upwellings of deep, cold water, with cold coastwise currents running toward the equator.
In North America, therefore, California and the Pacific coast of Mexico lack corals, while the West Indian region supports them in profusion. So do the coast of Brazil in South America, the tropical east African coast, and the northeastern shores of Australia, where the Great Barrier Reef creates a living wall for more than a thousand miles.
Within the United States the only coral coast is that of the Florida Keys. For nearly 200 miles these islands reach southwestward into tropical waters. They begin a little south of Miami where Sands, Elliott, and Old Rhodes Keys mark the entrance to Biscayne Bay; then other islands continue to the southwest, skirting the tip of the Florida mainland, from which they are separated by Florida Bay, and finally swinging out from the land to form a slender dividing line between the Gulf of Mexico and the Straits of Florida, through which the Gulf Stream pours its indigo flood.
To seaward of the Keys there is a shallow area three to seven miles wide where the sea bottom forms a gently sloping platform under depths generally less than five fathoms. An irregular channel (Hawk Channel) with depths to ten fathoms traverses these shallows and is navigable by small boats. A wall of living coral reefs forms the seaward boundary of the reef platform, standing on the edge of the deeper sea (see page 198).
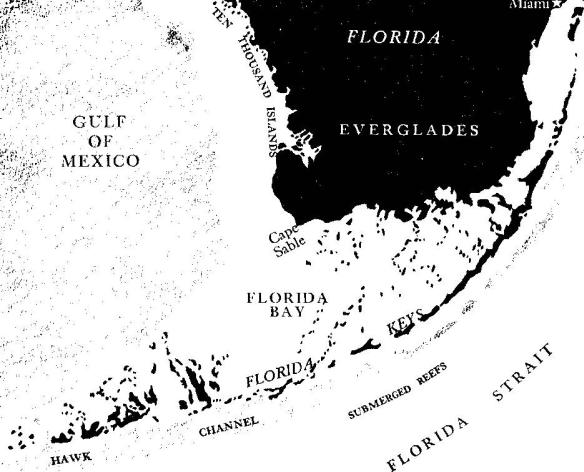
The Keys are divided into two groups that have a dual nature and origin. The eastern islands, swinging in their smooth arc 110 miles from Sands to Loggerhead Key, are the exposed remnants of a Pleistocene coral reef. Its builders lived and flourished in a warm sea just before the last of the glacial periods, but today the corals, or all that remains of them, are dry land. These eastern Keys are long, narrow islands covered with low trees and shrubs, bordered with coral limestone where they are exposed to the open sea, passing into the shallow waters of Florida Bay through a maze of mangrove swamps on the sheltered side. The western group, known as the Pine Islands, are a different kind of land, formed of limestone rock that had its origin on the bottom of a shallow interglacial sea, and is now raised only slightly above the surface of the water. But in all the Keys, whether built by the coral animals or formed of solidifying sea drift, the shaping hand is the hand of the sea.
In its being and its meaning, this coast represents not merely an uneasy equilibrium of land and water masses; it is eloquent of a continuing change now actually in progress, a change being brought about by the life processes of living things. Perhaps the sense of this comes most clearly to one standing on a bridge between the Keys, looking out over miles of water, dotted with mangrove‑covered islands to the horizon. This may seem a dreamy land, steeped in its past. But under the bridge a green mangrove seedling floats, long and slender, one end already beginning to show the development of roots, beginning to reach down through the water, ready to grasp and to root firmly in any muddy shoal that may lie across its path. Over the years the mangroves bridge the water gaps between the islands; they extend the mainland; they create new islands. And the currents that stream under the bridge, carrying the mangrove seedling, are one with the currents that carry plankton to the coral animals building the offshore reef, creating a wall of rocklike solidity, a wall that one day may be added to the mainland. So this coast is built.
To understand the living present, and the promise of the future, it is necessary to remember the past. During the Pleistocene, the earth experienced at least four glacial stages, when severe climates prevailed and immense sheets of ice crept southward. During each of these stages, large volumes of the earth’s water were frozen into ice, and sea level dropped all over the world. The glacial intervals were separated by milder interglacial stages when, with water from melting glaciers returning to the sea, the level of the world ocean rose again. Since the most recent Ice Age, known as the Wisconsin, the general trend of the earth’s climate has been toward a gradual, though not uniform warming up. The interglacial stage preceding the Wisconsin glaciation is known as the Sangamon, and with it the history of the Florida Keys is intimately linked.
The corals that now form the substance of the eastern Keys built their reef during that Sangamon interglacial period, probably only a few tens of thousands of years ago. Then the sea stood perhaps 100 feet higher than it does today, and covered all of the southern part of the Florida plateau. In the warm sea off the sloping southeastern edge of that plateau the corals began to grow, in water somewhat more than 100 feet deep. Later the sea level dropped about 30 feet (this was in the early stages of a new glaciation, when water drawn from the sea was falling as snow in the far north); then another 30 feet. In this shallower water the corals flourished even more luxuriantly and the reef grew upward, its structure mounting close to the sea surface. But the dropping sea level that at first favored the growth of the reef was to be its destruction, for as the ice increased in the north in the Wisconsin glacial stage, the ocean level fell so low that the reef was exposed and all its living coral animals were killed. Once again in its history the reef was submerged for a brief period, but this could not bring back the life that had created it. Later it emerged again and has remained above water, except for the lower portions, which now form the passes between the Keys. Where the old reef lies exposed, it is deeply corroded and dissected by the dissolving action of rain and the beating of salt spray; in many places the old coral heads are revealed, so distinctly that the species are identifiable.
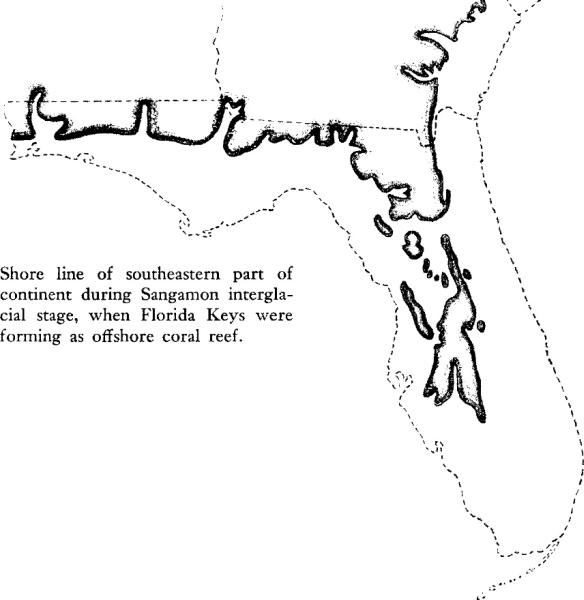
While the reef was a living thing, being built up in that Sangamon sea, the sediments that have more recently become the limestone of the western group of Keys were accumulating on the landward side of the reef. Then the nearest land lay 150 miles to the north, for all the southern end of the present Florida peninsula was submerged. The remains of many sea creatures, the solution of limestone rocks, and chemical reactions in the sea water contributed to the soft ooze that covered the shallow bottoms. With the changing sea levels that followed, this ooze became compacted and solidified into a white, fine‑textured limestone, containing many small spherules of calcium carbonate resembling the roe of fish; because of this characteristic it is sometimes known as “oolitic limestone,” or “Miami oolite.” This is the rock immediately underlying the southern part of the Florida mainland. It forms the bed of Florida Bay under the layer of recent sediments, and then rises above the surface in the Pine Islands, or western Keys, from Big Pine Key to Key West. On the mainland, the cities of Palm Beach, Fort Lauderdale, and Miami stand on a ridge of this limestone formed when currents swept past an old shore line of the peninsula, molding the soft oozes into a curving bar. The Miami oolite is exposed on the floor of the Everglades as rock of strangely uneven surface, here rising in sharp peaks, there dropping away in solution holes. Builders of the Tamiami Trail and of the highway from Miami to Key Largo dredged up this limestone along the rights of way and with it built the foundations on which these highways are laid.
Knowing this past, we can see in the present a repetition of the pattern, a recurrence of earth processes of an earlier day. Now, as then, living reefs are building up offshore; sediments are accumulating in shallow waters; and the level of the sea, almost imperceptibly but certainly, is changing.
Off this coral coast the sea lies green in the shallows, blue in the far distances. After a storm, or even after a prolonged southeasterly blow, comes “white water.” Then a thick, milk‑white, richly calcareous sediment is washed out of the reefs and stirred from its deep beds over the floor of the reef flat. On such days the diving mask and the aqualung may as well be left behind, for the underwater visibility is little better than in a London fog.

“White water” is the indirect result of the very high rate of sedimentation that prevails in the shallows around the Keys. Anyone who wades out even a few steps from the shore notices the white, siltlike substance adrift in the water and accumulating on the bottom. It has visibly rained down on every surface. Its fine dust lies over sponge and gorgonian and anemone; it chokes and buries the low‑growing algae and lies whitely over the dark bulks of the big loggerhead sponges. The wader stirs up clouds of it; winds and strong currents set it in motion. Its accumulation is going on at an astonishing rate; sometimes, after a storm, two or three inches of new sediment are deposited from one high tide to the next. It comes from various sources. Some is mechanically derived from the disintegration of dead plants and animals–mollusk shells, lime‑depositing algae, coral skeletons, tubes of worms or snails, spicules of gorgonians and sponges, skeletal plates of holothurians. It is also derived in part from chemical precipitation of the calcium carbonate present in the water. This, in turn, has been leached out of the vast expanses of limestone rock that compose the surface of southern Florida, and has been carried to the sea by rivers and by the slow drainage of the Everglades.
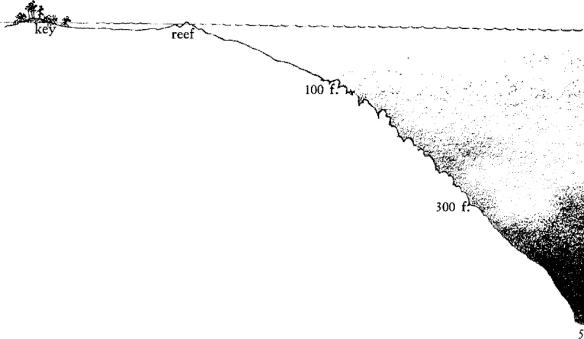
A few miles outside the chain of the present Keys is the reef 5 of living coral, forming the seaward rim of the shallows, and overlooking a steep descent into the trough of the Florida straits. The reefs extend from Fowey Rocks, south of Miami, to the Marquesas and Tortugas and in general they mark the 10‑fathom depth contour. But often they rise to lesser depths and here and there they break the surface as tiny offshore islands, many of them marked by lighthouses.
Drifting over the reef in a small boat and peering down through a glass‑bottomed bucket, one finds it hard to visualize the whole terrain, for so little of it can be seen at a time. Even a diver exploring more intimately finds it difficult to realize he is on the crest of a high hill, swept by currents instead of winds, where gorgonians are the shrubbery and stands of elkhorn coral are trees of stone. Toward the land, the sea floor slopes gently down from this hilltop into the wide water‑filled valley of Hawk Channel; then it rises again and breaks water as a chain of low‑lying islands–the Keys. But on the seaward side of the reef the bottom descends quickly into blue depths. Live corals grow down to a depth of about 10 fathoms. Below that it is too dark, perhaps, or there is too much sediment, and instead of living coral there is a foundation of dead reef, formed at some time when the sea level was lower than it is today. Out where the water is about 100 fathoms deep there is a clean rock bottom, the Pourtal[[[grave.gif]]]s Plateau; its fauna is rich, but the corals that live here are not reef builders. Between 300 and 500 fathoms sediments have again accumulated on a slope that descends to the trough of the Florida straits–the channel of the Gulf Stream.
As for the reef itself, many thousand thousand beings–plant and animal, living and dead–have entered into its composition. Corals of many species, building their little cups of lime and with them fashioning many strange and beautiful forms, are the foundation of the reef. But besides the corals there are other builders and all the interstices of the reef are filled with their shells or their limy tubes, or with coral rock cemented together with building stones of the most diverse origin. There are colonies of tube‑building worms and there are mollusks of the snail tribe whose contorted, tubular shells may be intertwined into massive structures. Calcareous algae, which have the property of depositing lime in their living tissues, form part of the reef itself or, growing abundantly over the shallows on the landward side, add their substance at death to the coral sand of which limestone rock is later formed. The horny corals or gorgonians, known as sea fans and sea whips, all contain limestone spicules in their soft tissues. These, along with lime from starfish and sea urchins and sponges and an immense number of smaller creatures, will eventually, with the passage of time and through the chemistry of the sea, come to form part of the reef.
Along with those that build are others that destroy. The sulphur sponge dissolves away the calcareous rock. Boring mollusks riddle it with their tunnels, and worms with sharp, biting jaws eat into it, weakening its structure and so hastening the day when a mass of coral will yield to the force of the waves, will break away, and perhaps roll down the seaward face of the reef into deeper water.
The basis of this whole complex association is a minute creature of deceptively simple appearance, the coral polyp. The coral animal is formed on the same general lines as the sea anemone. It is a double‑walled tube of cylindrical shape, closed at the base and open at the free end, where a crown of tentacles surrounds the mouth. The important difference–the fact on which the existence of coral reefs depends–is this: the coral polyp has the ability to secrete lime, forming a hard cup about itself. This is done by cells of the outer layer, much as the shell of a mollusk is secreted by an outer layer of soft tissue–the mantle. So the anemone‑like coral polyp comes to sit in a compartment formed of a substance as hard as rock. Because the “skin” of the polyp is turned inward at intervals in a series of vertical folds, and because all of this skin is actively secreting lime, the cup does not have a smooth circumference, but is marked by partitions projecting inward, forming the starlike or flowerlike pattern familiar to anyone who has examined a coral skeleton.
Most corals build colonies of many individuals. All the individuals of any one colony, however, are derived from a single fertilized ovum that matured and then began to form new polyps by budding. The colony has a shape characteristic of the species–branched, boulderlike, flatly encrusting, or cup‑shaped. Its core is solid, for only the surface is occupied by living polyps, which may be widely separated in some species or closely crowded in others. It is often true that the larger and more massive the colony, the smaller the individuals that compose it; the polyps of a branching coral taller than a man may themselves be only an eighth of an inch high.

The hard substance of the coral colony is usually white, but may take on the colors of minute plant cells that live within the soft tissues in a relation of mutual benefit. There is the exchange usual in such relations, the plants getting carbon dioxide and the animals making use of the oxygen given off by the plants. This particular association may have a deeper significance, however. The yellow, green, or brown pigments of the algae belong to the group of chemical substances known as carotinoids. Recent studies suggest that these pigments in the imprisoned algae may act on the corals, serving as “internal correlators” to influence the processes of reproduction. Under normal conditions, the presence of the algae seems to benefit the coral, but in dim light the coral animals rid themselves of the algae by excreting them. Perhaps this means that in weak light or in darkness the whole physiology of the plant is changed and the products of its metabolism are altered to something harmful, so that the animal must expel the plant guest.
Within the coral community there are other strange associations. In the Florida Keys and elsewhere in the West Indian region, a gall crab makes an oven‑shaped cavity on the upper surface of a colony of living brain coral. As the coral grows the crab manages to keep open a semicircular entrance through which, while young, it enters and leaves its den. Once full grown, however, the crab is believed to be imprisoned within the coral. Few details of the existence of this Florida gall crab are known, but in a related species in corals of the Great Barrier Reef only the females form galls. The males are minute, and apparently visit the females in the cavities where they are imprisoned. The female of this species depends on straining food organisms from indrawn currents of sea water and its digestive apparatus and appendages are much modified.

Everywhere, throughout the whole structure of the reef as well as inshore, the horny corals or gorgonians are abundant, sometimes outnumbering the corals. The violet‑hued sea fan spreads its lace to the passing currents, and from all the structure of the fan innumerable mouths protrude through tiny pores, and tentacles reach out into the water to capture food. The little snail known as the flamingo tongue, wearing a solid and highly polished shell, often lives on the sea fans. The soft mantle, extended to cover the shell, is a pale flesh color with numerous black, roughly triangular markings. The gorgonians known as sea whips are more abundant, forming dense stands of undersea shrubbery, often waist‑high, sometimes as tall as a man. Lilac, purple, yellow, orange, brown, and buff are the colors worn by these gorgonians of the coral reefs.
Encrusting sponges spread their mats of yellow, green, purple, and red over the walls of the reef; exotic mollusks like the jewel box and the spiny oyster cling to it; long‑spined sea urchins make dark, bristling patches in the hollows and crevices; and schools of brightly colored fishes twinkle along the façade of the reef where the lone hunters, the gray snapper and the barracuda, wait to seize them.
At night the reef comes alive. From every stony branch and tower and domed façade, the little coral animals, who, avoiding daylight, had remained shrunken within their protective cups until darkness fell, now thrust out their tentacled heads and feed on the plankton that is rising toward the surface. Small crustacea and many other forms of microplankton, drifting or swimming against a branch of coral, are instant victims of the myriad stinging cells with which each tentacle is armed. Minute though the individual plankton animals be, the chances of passing unharmed through the interlacing branches of a stand of elkhorn coral seem slender indeed.
Other creatures of the reef respond to night and darkness and many of them emerge from the grottoes and crevices that served as daytime shelter. Even that strange hidden fauna of the massive sponges–the small shrimps and amphipods and other animals that live as unbidden guests deep within the canals of the sponge–at night creep up along those dark and narrow galleries and collect near their thresholds as though looking out upon the world of the reef.
On certain nights of the year, extraordinary events occur over the reefs. The famed palolo worm of the South Pacific, moved to gather in its prodigious spawning swarms on a certain moon of a certain month–and then only–has its less‑known counterpart in a related worm that lives in the reefs of the West Indies and at least locally in the Florida Keys. The spawning of this Atlantic palolo has been observed repeatedly about the Dry Tortugas reefs, at Cape Florida, and in several West Indian localities. At Tortugas it takes place always in July, usually when the moon reaches its third quarter, though less often on the first quarter. The worms never spawn on the new moon.

The palolo inhabits burrows in dead coral rock, sometimes appropriating the tunnelings of other creatures, sometimes excavating its burrow by biting away fragments of rock. The life of this strange little creature seems to be ruled by light. In its immaturity the palolo is repelled by light–by sunlight, by the light of the full moon, even by paler moonlight. Only in the darkest hours of the night, when this strong inhibition of the light rays is removed, does it venture from its burrow, creeping out a few inches in order to nibble at the vegetation on the rocks. Then, as the season for spawning approaches, remarkable changes take place within the bodies of the worms. With the maturing of the sex cells, the segments of the posterior third of each animal take on a new color, deep pink in the males, greenish gray in the females. Moreover, this part of the body, distended with eggs or sperm, becomes exceedingly thin‑walled and fragile, and a noticeable constriction develops between this and the anterior part of the worm.
At last there comes a night when these worms–so changed in their physical beings–respond in a new way to the light of the moon. No longer does the light repel and hold them prisoners within their burrows. Instead, it draws them out to the performance of a strange ritual. The worms back out of their burrows, thrusting out the swollen, thin‑walled posterior ends, which immediately begin a series of twisting movements, writhing in spiral motions until suddenly the body breaks at the weak point and each worm becomes two. The two parts have different destinies–the one to remain behind in the burrow and resume the life of the timid forager of the dark hours, the other to swim up toward the surface of the sea, to become one of a vast swarm of thousands upon thousands of worms joining in the spawning activities of the species.
During the last hours of the night the number of swarming worms increases rapidly, and when dawn comes the sea over the reef is almost literally filled with them. When the first rays of the sun appear, the worms, strongly stimulated by the light, begin to twist and contract violently, their thin‑walled bodies burst open, and the eggs from some and sperm from others are cast into the sea. The spent and empty worms may continue to swim weakly for a short time, preyed upon by fish that gather for a feast, but soon all that remain have sunk to the bottom and died. But floating at the surface of the sea are the fertilized eggs, drifting over areas many feet deep and acres in extent. Within them swift changes have begun–the division of cells, the differentiation of structure. By evening of that same day the eggs have yielded up tiny larvae, swimming with spiral motions through the sea. For about three days the larvae live at the surface; then they become burrowers in the reefs below until, a year hence, they will repeat the spawning behavior of their kind.

Some related worms that swarm periodically about the Keys and the West Indies are luminous, creating beautiful pyrotechnic displays on dark nights. Some people believe that the mysterious light reported by Columbus as seen by him on the night of October 11, “about four hours before making the landfall and an hour before moonrise,” may have been a display of some of these “fireworms.”
The tides pouring in from the reefs and sweeping over the flats come to rest against the elevated coral rock of the shore. On some of the Keys the rock is smoothly weathered, with flattened surfaces and rounded contours, but on many others the erosive action of the sea has produced a rough and deeply pitted surface, reflecting the solvent action of centuries of waves and driven salt spray. It is almost like a stormy sea frozen into solidity, or as the surface of the moon might be. Little caves and solution holes extend above and below the line of the high tide. In such a place I am always strongly aware of the old, dead reef beneath my feet, and of the corals whose patterns, now crumbling and blurred, were once the delicately sculptured vessels that held the living creatures. All the builders now are dead–they have been dead for thousands of years–but that which they created remains, a part of the living present.
Crouching on the jagged rocks, I hear little murmurings and whisperings born of the movements of air and water over these surfaces–the audible voice of this nonhuman, intertidal world. There are few obvious signs of life to break the spell of brooding desolation. Perhaps a dark‑bodied isopod–a sea roach–darts across the dry rock to disappear into one of the small sea caves, daring exposure to light and to sharp‑eyed enemies only for the moment of its swift passage from one dark recess to another. There are thousands of its kind in the coral rock, but not until darkness covers the shore will they come out in numbers to search for the bits of animal and vegetable refuse that are their food.
At the high‑tide line, growths of microscopic plants darken the coral rock, tracing that mysterious black line that marks the sea’s edge on all rocky coasts of the world. Because of the irregular surface and deep dissection of the coral rock, the sea runs in under the high‑tide rocks by way of crevices and depressions, and so the black zone darkens the jagged peaks and the rims of holes and little caves, while lighter rock of a yellowish‑gray hue lines the depressions below that controlling tidal level.

Small snails whose shells are boldly striped or checked in black and white–the neritas–crowd down into cracks and cavities in the coral or rest on open rock surfaces waiting for the return of the tide when they can feed. Others, in rounded shells with roughly beaded surfaces, belong to the periwinkle tribe. Like many others of their kind, these beaded periwinkles are making a tentative invasion of the land, living under rocks or logs high on the shore or even entering the fringe of land vegetation. Black horn shells live in numbers just below the line of the high tides, feeding on the algal film over the rocks. The living snails are held by some intangible bonds to this tidal level, but the shells discarded after their death are found and taken as habitations by the smallest of the hermit crabs, who then carry them down onto the lower levels of the shore.
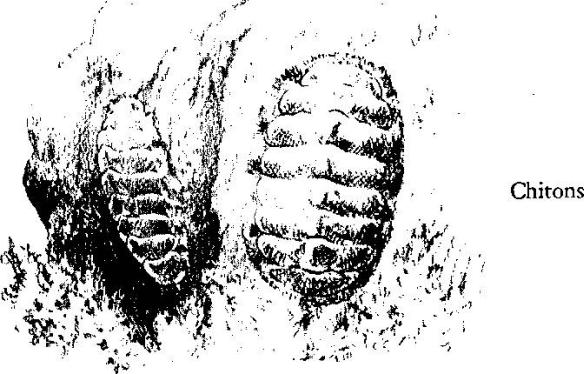
These deeply eroded rocks are the home of the chitons, whose primitive form harks back to some ancient group of mollusks of which they are the only living representatives. Their oval bodies, covered with a jointed shell of eight transverse plates, fit into depressions in the rocks when the tide is out. They grip the rocks so strongly that even heavy waves can get no hold on their sloping contours. When the high tide covers them, they begin to creep about, resuming their rasping of vegetation from the rocks, their bodies swaying to and fro in time to the scraping motions of the radula or file‑like tongue. Month in and month out, a chiton moves only a few feet in any direction; because of this sedentary habit, the spores of algae and the larvae of barnacles and tube‑building worms settle upon its shell and develop there. Sometimes, in dark wet caves, the chitons pile up, one on top of another, and each scrapes algae off the back of the one beneath it. In a small way these primitive mollusks may be an agent of geologic change as they feed on the rocks, each removing, along with the algae, minute scrapings of rock particles and so, over the centuries and the millennia in which this ancient race of beings has lived its simple life, contributing to the processes of erosion by which earth surfaces are worn away.
On some of these Keys a small intertidal mollusk called Onchidium lives deep in little rock caverns, the entrances of which are often overgrown by colonies of mussels. Although it is a mollusk and a snail, Onchidium has no shell. It belongs to a group that consists largely of land snails or slugs, in many of which the shell is lacking or concealed. Onchidium inhabits tropical seashores, living usually on beaches of roughly eroded rock. As the tide falls, processions of small black slugs emerge from their doorways, wriggling and pushing their way out through the impeding mussel threads, a dozen or more individuals coming out of a common cave to feed on the rocks, from which they scrape vegetation as the chitons do. As they emerge, each is invested with a tunic of slime that makes it look jet black, wet, and shining; in wind and sun the little slug dries to a deep blue‑black, over which is a slight, milky bloom.
On these journeys the slugs seem to follow haphazard or irregular paths over the rocks. They continue feeding as the tide falls to its lowest ebb, and even as it turns and begins to rise. About half an hour before the returning sea has reached them, and before so much as a drop of water has splashed into their nests, all of the slugs cease their grazing and begin to return to the home nest. While the outgoing path was meandering, the return is by a direct route. The members of each community return to their own nest, even though the way may lie over greatly eroded rock surfaces and even though the path may cross the routes of other slugs returning to other nests. All of the individuals belonging to one nest‑community, even though they may have been widely separated while feeding, begin the return journey at almost the same moment. What is the stimulus? It is not the returning water, for that has not touched them; when it laps again over their rocks they will be safe within their nests.
The whole pattern of behavior of this little creature is puzzling. Why should it be drawn to live again at the edge of the sea that its ancestors deserted thousands or millions of years ago? It comes forth only when the tide has fallen, then, somehow sensing the impending return of the sea and seeming to remember its recent affinities with the land, it hurries to safety before the tide can find it and carry it away. How has it acquired this behavior, attracted yet repelled by the sea? We can only ask these questions; we cannot answer them.
For its protection during the feeding journeys, Onchidium is equipped with means of detecting and driving away its enemies. Minute papillae on its back are sensitive to light and passing shadows. Other, stouter papillae associated with the mantle are equipped with glands that secrete a milky, highly acid fluid. If the animal is suddenly disturbed, it expels spurting streams of this acid, the streams breaking up in the air to a fine spray that may be thrown five or six inches, or as much as a dozen times the length of the animal. The old German zoologist Semper, who studied a species of Onchidium in the Philippines, believed this dual equipment served to protect the slug from the beach‑hopping blenny, a fish of many tropical mangrove coasts that leaps along above the tide, feeding on Onchidium and crabs. Semper thought the slugs could detect the shadow of an approaching fish and drive off the enemy by discharging the white acid spray. In Florida or elsewhere in the West Indian region there is no fish that comes out of water to pursue its prey. On the rocks where Onchidium must feed there are, however, scrambling crabs and isopods whose jostlings might well push the slugs into the water, for they have no means of gripping the rocks. For whatever reason, the slugs react to the crabs and to the isopods as to dangerous enemies, responding to their touch by discharging the repellant chemical.
In the strip between tropical tide lines, conditions are difficult for nearly all forms of life. The heat of the sun increases the hazards of exposure during the withdrawal of the tide. The shifting layers of choking sediment, accumulating on flat or gently sloping surfaces, discourage many plants and animals of types that inhabit rocky shores in the clearer, cooler waters of the north. Instead of the vast barnacle and mussel fields of New England there are only scattering patches of these creatures, varying from Key to Key but never really abundant. Instead of the great rockweed forests of the north, there are only scattered growths of small algae, including various brittle, lime‑secreting forms, none of which offer shelter or security to any considerable number of animals.
If the area marked out by the advance and retreat of the neap tides is in general inhospitable, there are nevertheless two forms of life–one plant, one animal–that are thoroughly at home there, and live in profusion nowhere else. The plant is a peculiarly‑beautiful alga that resembles spheres of green glass clustered together in irregular masses. It is Valonia, the sea bottle, a green alga that forms large vesicles filled with a sap that bears a definite chemical relation to the water about it, varying the proportions of its contained ions of sodium and potassium according to variations in the intensity of sunlight, the exposure to surf, and other conditions of its world. Under overhanging rock and in other sheltered places it forms sheets and masses of its emerald globules, lying half buried in deep drifts of sediment.

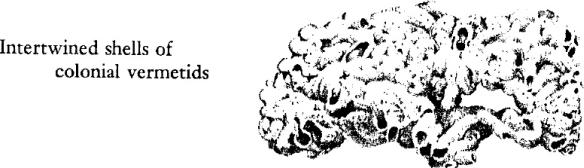
The animal symbol of this intertidal world of coral is a group of snails whose whole structure and being represent an extraordinary contrast to the way of life typical of this class of mollusks. They are called the vermetid or “wormlike” snails. The shell is no ordinary gastropod spire or cone, but a loose uncoiled tube very like the calcareous tubes built by many worms. The species that inhabit this intertidal zone have become colonial, and their tubes form closely packed and intertwined masses.
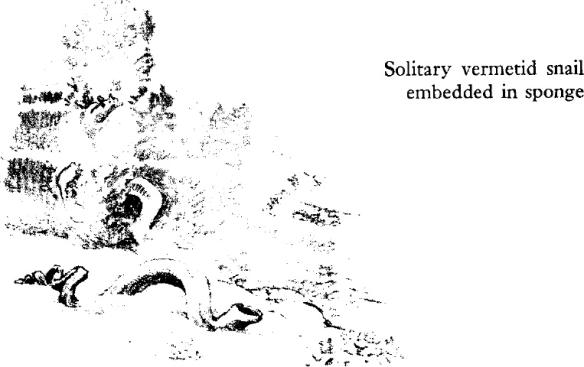
The very nature of these vermetid snails and their departure from the form and habits of related mollusks are eloquent of the circumstances of their world and of the readiness of life to adapt itself to a vacant niche. Here on this coral platform the tide ebbs and flows twice daily, and each flood brings renewed food supplies from offshore. There is but one perfect way to exploit such rich supplies: to remain in one place and fish the currents as they stream by. This is done on other shores by such animals as the barnacles, the mussels, and the tube‑building worms. It is not ordinarily a snail’s way of life, but in adaptation these extraordinary snails have become sedentary, abandoning the typical roaming habit. No longer solitary, they have become gregarious to an extreme degree, living in crowded colonies, with shells so intertwined that early geologists called their formations “worm rock.” And they have given up the snail habits of scraping food from the rocks or of hunting and devouring other animals of large size; instead they draw the sea water into their bodies and strain out the minute planktonic food organisms. The tips of their gills are thrust out and drawn through the water like nets–an adaptation probably unique in all the group of snail‑like mollusks. The vermetids give their own clear demonstration of the plasticity of the living organism and its responsiveness to the world about it. Again and again, in group after group of widely different and unrelated animals, the same problem has been met and solved by the evolution of diverse structures that function for a common purpose. So the legions of the barnacles sweep food from the tides on a New England shore, using a modification of what in their relatives would be a swimming appendage; mole crabs gather by the thousand where surf sweeps the southern beaches, straining out food with the bristles of their antennae; and here on this coral shore the crowded masses of this strange snail filter the waters of the incoming tide through their gills. By becoming the imperfect, the atypical snail, they have become the perfectly adapted exploiter of the opportunities of their world.
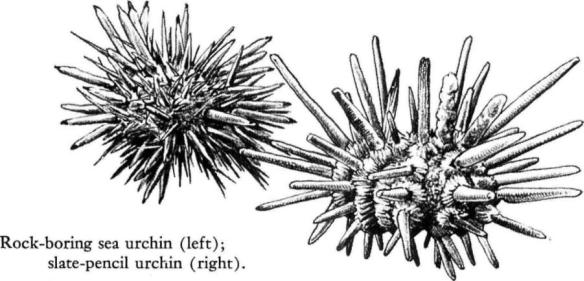
The edge of the low tide is a dark line traced by colonies of short‑spined, rock‑boring sea urchins. Every hole and every depression in the coral rock bristles with their small dark bodies. One spot in the Keys lives in my memory as an urchin paradise. This is the seaward shore of one of the eastern group of islands, where the rock drops in an abrupt terrace, somewhat undercut and deeply eroded into holes and small caves, many with their roofs open to the sky. I have stood on the dry rock above the tide and looked down into these little water‑floored, rock‑walled grottoes, finding twenty‑five to thirty urchins in one of these caverns that was no larger than a bushel basket. The caves shine with a green water‑light in the sun, and in this light the globular bodies of the urchins have a reddish color of glowing, luminous quality, in rich contrast to the black spines.

A little beyond this spot the sea bottom slopes under water more gradually, with no undercutting. Here the rock borers seem to have taken over every niche that can afford shelter; they give the illusion of shadows beside each small irregularity of bottom. It is not certain whether they use the five short stout teeth on their under surfaces to scrape out holes in the rock, or perhaps merely take advantage of natural depressions to find a safe anchorage against the occasional storms that sweep this coast. For some inscrutable reason, these rock‑boring urchins and related species in other parts of the world are bound to this particular tidal level, linked to it precisely and mysteriously by invisible ties that prevent their wandering farther out over the reef flat, although other species of urchins are abundant there.
Above and below the zone of the rock‑boring urchins, closely crowded throngs of pale brown tubular creatures push up through the chalky sediment. When the tide leaves them their tissues retract and all that proclaims them to be animals is hidden; then one might pass them by as some strange marine fungi. With the return of the water their animal nature is revealed, and from each fawn‑colored tube a crown of tentacles, of purest emerald green, is unfolded as each of these anemone‑like creatures begins to search the tide for the food it has brought. Living where their very existence depends on keeping the delicate tissues of the tentacles above the choking dust of sediment, these zoanthids are able to stretch their bodies into slender threads where the sediments are deep, though normally their tubes are short and stout.
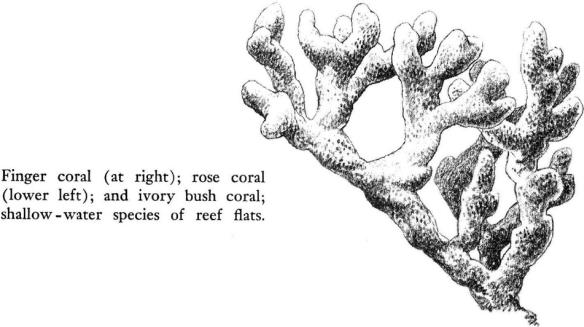
On the seaward side of many of the Keys the bottom slopes gently, with wading depths for perhaps a quarter of a mile or more. Once beyond the rock‑boring sea urchins, the vermetid snails, and the green and brown jewel anemones, the bottom of coarse sand and coral fragments begins to be marked by dark patches of turtle grass, and larger animals begin to inhabit the reef flats. Sponges, dark and bulky, grow in water only deep enough to cover their massive forms. Small, shallow‑water corals, somehow able to survive the rain of sediments that would be fatal to the larger reef‑builders, erect their hard structures, stoutly branched or domed, on the floor of coral rock. The gorgonians, plantlike in their habit of growth, are a low shrubbery of delicate rose and brown and purple hues. And within and among and beneath them all is the infinitely varied fauna of a tropical coast, as many creatures that wander freely through the waters of this warm sea crawl or swim or glide over the flats.
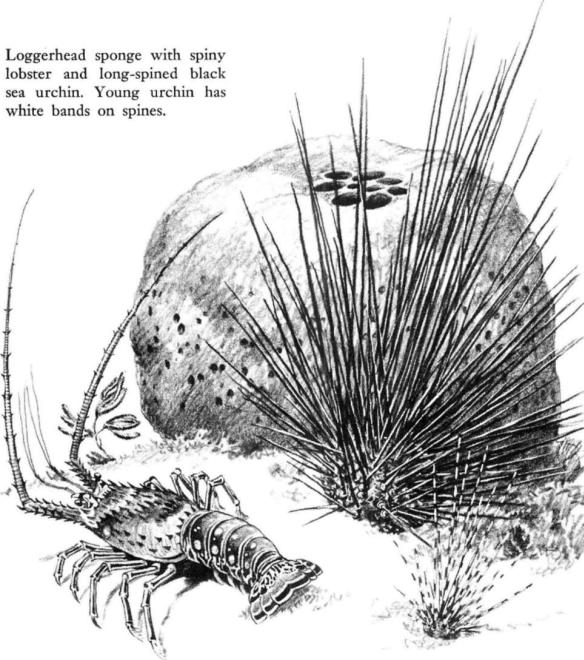
Massive and inert, the loggerhead sponges by their appearance suggest nothing of the activity that goes on within their dark bulks. There is no sign of life for the casual passer‑by to read, although if he waited and watched long enough he might sometimes see the deliberate closing of some of the round openings, large enough to admit an exploring finger, that penetrate the flat upper surface. These and other openings are the key to the nature of the giant sponge which, like even the smallest of its group, can exist only as long as it can keep the waters of the sea circulating through its body. Its vertical walls are pierced by intake canals of small diameter, groups of them covered by sieve plates with numerous perforations. From these the canals lead almost horizontally into the interior of the sponge, branching and rebranching into tubes of progressively smaller bore, to penetrate all the massive bulk of the sponge and finally to lead up to the large exit canals. Perhaps these exit holes are kept free of choking sediment by the strength of the outbound currents; at any rate they are the only part of the sponge that shows a pure black color, for the flour‑like whiteness of the reef sediments has been sifted over all the sooty black surface of the body.
In its passage through the sponge, the water leaves a coating of minute food organisms and organic detritus on the walls of the canals; the cells of the sponge pick up the food, pass the digestible materials along from cell to cell, and return waste material to the flowing currents. Oxygen passes into the sponge cells; carbon dioxide is given off. And sometimes small sponge larvae, having undergone the early stages of their development within the parent sponge, detach themselves and enter the dark flowing river, to pass with it into the sea.
The intricate passageways, the shelter and available food they offer, have attracted many small creatures to live within the sponge. Some come and go; others never leave the sponge once they have taken up residence within it. One such permanent lodger is a small shrimp–one of the group known as snapping shrimp because of the sound made by snapping the large claw. Although the adults are imprisoned, the young shrimp, hatched from eggs adhering to the appendages of their mothers, pass out with the water currents into the sea and live for a time in the currents and tides, drifting, swimming, perhaps carried far afield. By mischance they may occasionally find their way into deep water where no sponges grow. But many of the young shrimp will in time find and approach the dark bulk of some loggerhead sponge and, entering it, will take up the strange life of their parents. Wandering through its dark halls, they scrape food from the walls of the sponge. As they creep along these cylindrical passageways, they carry their antennae and their large claws extended before them, as though to sense the approach of a larger and possibly dangerous creature, for the sponge has many lodgers of many species–other shrimps, amphipods, worms, isopods–and their numbers may reach into the thousands if the sponge is large.
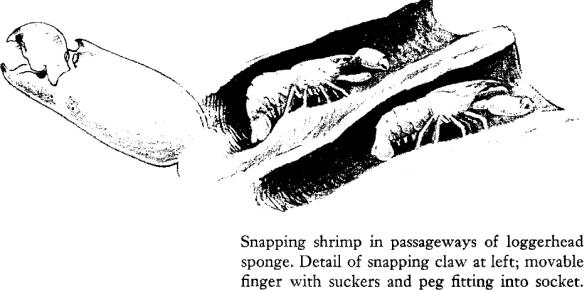
There, on the flats off some of the Keys, I have opened small loggerheads and heard the warning snapping of claws as the resident shrimps, small, amber‑colored beings, hurried into the deeper cavities. I had heard the same sound filling the air about me, as, on an evening low tide, I waded in to the shore. From all the exposed reef rock there were strange little knockings and hammerings, yet the sounds, to a maddening degree, were impossible to locate. Surely this nearby hammering came from this particular bit of rock; yet when I knelt to examine it closely there was silence; then from all around, from everywhere but this bit of rock at hand, all the elfin hammering was resumed. I could never find the little shrimps in the rocks, yet I knew they were related to those I had seen in the loggerhead sponges. Each has one immense hammer claw almost as long as the rest of its body. The movable finger of the claw bears a peg that fits into a socket in the rigid finger. Apparently the movable finger, when raised, is held in position by suction. To lower it, extra muscular force must be applied, and when the suction is overcome, it snaps into place with audible sound, at the same time ejecting a spurt of water from its socket. Perhaps the water jet repels enemies and aids in capturing prey, which may also be stunned by a blow from the forcibly retracted claw. Whatever the value of the mechanism, the snapping shrimps are so abundant in the shallows of tropical and subtropical regions, and snap their claws so incessantly, that they are responsible for much of the extraneous noise picked up on underwater listening devices, filling the water world with a continuous sizzling, crackling sound.
It was on the reef flats off Ohio Key, on a day early in May, that I had my first, startled encounter with tropical sea hares. I was wading over a part of the flat that had an unusually heavy growth of rather tall seaweeds when sudden movement drew my eyes to several heavy‑bodied, foot‑long animals moving among the weeds. They were a pale tan color, marked with black rings, and when I touched one cautiously with my foot, it responded instantly by expelling a concealing cloud of fluid the color of cranberry juice.
I had met my first sea hare years before on the North Carolina coast. It was a small creature about as long as my little finger, browsing peacefully among some seaweeds near a stone jetty. I slipped my hand under it and gently brought it toward me, then, its identity confirmed, I returned the little creature carefully to the algae, where it resumed its grazing. Only by drastic revision of my mental image could I accept these tropical creatures, which seemed to belong in some book of mythology, as relatives of that first little elfin being.
The large West Indian sea hares inhabit the Florida Keys as well as the Bahamas, Bermuda, and the Cape Verde Islands. Within their range they usually live offshore, but at the spawning season they move in to the shallows, where I had found them, to attach their eggs, in tangled threads, to the weeds near the low‑tide mark. They are marine snails of a sort, but have lost their external shells and possess only an internal remnant, hidden by the soft mantle tissue. Two prominent tentacles suggestive of ears, and the rabbit‑like body shape, are responsible for the common name (see page 235).
Whether because of its strange appearance, or because of its defensive fluids, often thought to be poisonous, the Old World sea hare has long had a secure place in folk lore, superstition, and witchcraft. Pliny declared it was poisonous to the touch, and recommended as an antidote asses’ milk and ground asses’ bones, boiled together. Apuleius, known chiefly as the author of The Golden Ass, became curious about the internal anatomy of the sea hare and persuaded two fishermen to bring him a specimen; whereupon he was accused of witchcraft and poisoning. Some fifteen centuries were to pass before anyone else ventured to publish a description of the internal anatomy of the creature–then Redi in 1684 described it, and although popular belief called it sometimes a worm, sometimes a holothurian, sometimes a fish, he placed it correctly, at least as to general relationships, as a marine slug. For the past century or more the harmless nature of the sea hares has been recognized for the most part, but although they are fairly well known in Europe and Great Britain, the American sea hares, largely confined to tropical waters, are less familiar animals.
Perhaps this anonymity is due in part to the infrequency of their spawning migrations into tidal waters. An individual animal is both male and female; it may function as either sex, or as both. In laying its eggs, the sea hare extrudes a long thread in little spurts, about an inch at a time, continuing the slow process until the string has reached a length sometimes as great as 65 feet, and contains about 100,000 eggs. As the pink or orange‑colored thread is expelled it curls about the surrounding vegetation, forming a tangled mass of spawn. The eggs and the resulting young meet the common fate of marine creatures; many eggs are destroyed, being eaten by crustacea or other predators (even by their own kind), and many of the hatching larvae fail to survive the dangers of life in the plankton. In the drift of the currents the larvae are carried offshore, and when they undergo metamorphosis to the adult form and seek the bottom they are in deep water. Their color changes with changing food as they migrate shoreward: first they are a deep rose color, then they are brown, then olive‑green like the adults. For one of the European species, at least, the known life history suggests a curious parallel with that of the Pacific salmon. With maturity, the sea hares turn shoreward to spawn. It is a journey from which there is no return; they do not reappear on the offshore feeding grounds, but apparently die after this single spawning.
The world of the reef flats is inhabited by echinoderms of every sort: starfishes, brittle stars, sea urchins, sand dollars, and holothurians all are at home on the coral rock, in the shifting coral sands, among the gorgonian sea gardens and the grass‑carpeted bottoms. All are important in the economy of the marine world–as links in the living chains by which materials are taken from the sea, passed from one to another, returned to the sea, borrowed again. Some are important also in the geologic processes of earth building and earth destruction–the processes by which rock is worn away and ground to sand, by which the sediments that carpet the sea floor are accumulated, shifted, sorted, and distributed. And at death their hard skeletons contribute calcium for the needs of other animals or for the building of the reefs.
Out on the reefs the long‑spined black sea urchin excavates cavities along the base of the coral wall; each sinks into its depression and turns its spines outward, so that a swimmer moving along the reef sees forests of black quills. This urchin also wanders in over the reef flats, where it nestles close to the base of a loggerhead sponge, or sometimes, apparently finding no need of concealment, rests in open, sand‑floored areas.
A full‑grown black urchin may have a body or test nearly 4 inches in diameter, with spines 12 to 15 inches long. This is one of the comparatively few shore animals that are poisonous to the touch, and the effect of contact with one of the slender, hollow spines is said to be like that of a hornet sting, or may even be more serious for a child or an especially susceptible adult. Apparently the mucous coating of the spines bears the irritant or poison.
This urchin is extraordinary in the degree of its awareness of the surroundings. A hand extended over it will cause all the spines to swivel about on their mountings, pointing menacingly at the intruding object. If the hand is moved from side to side the spines swing about, following it. According to Professor Norman Millott of the University College of the West Indies, nerve receptors scattered widely over the body receive the message conveyed by a change in the intensity of light, responding most sharply to suddenly decreased light as a shadowy portent of danger. To this extent, then, the urchin may actually “see” moving objects passing nearby.
Linked in some mysterious way with one of the great rhythms of nature, this sea urchin spawns at the time of the full moon. The eggs and sperm are shed into the water once in each lunar month during the summer season, on the nights of strongest moonlight. Whatever the stimulus to which all the individuals of the species respond, it assures that prodigal and simultaneous release of reproductive cells that nature often demands for the perpetuation of a species.
Off some of the Keys, in shallow water, lives the so‑called slate‑pencil urchin, named for its short stout spines. This is an urchin of solitary habit, single individuals sheltering under or among the reef rocks near the low‑tide level. It seems a sluggish creature of dull perceptions, unaware of the presence of an intruder, and making no effort to cling by means of its tube feet when it is picked up. It belongs to the only family of modern echinoderms that also existed in Paleozoic time; the recent members of the group show little change from the form of ancestors that lived hundreds of millions of years ago.
Another urchin with short and slender spines and color variations ranging from deep violet to green, rose, or white, sometimes occurs abundantly on sandy bottoms carpeted with turtle grass, camouflaging itself with bits of grass and shell and coral fragments held in its tube feet. Like many other urchins, it performs a geologic function. Nibbling away at shells and coral rock with its white teeth, it chips off fragments that are then passed through the grinding mill of its digestive tract; these organic fragments, trimmed, ground, and polished within the urchins, contribute to the sands of tropical beaches.
And the tribes of the starfish and the brittle stars are everywhere represented on these coral flats. The great sea star, Oreaster, stout and powerful of body, perhaps lives more abundantly a little offshore, where whole constellations of them gather on the white sand. But solitary specimens wander inshore, seeking especially the grassy areas.
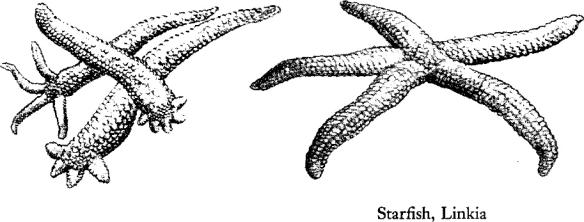
A small reddish‑brown starfish, Linkia, has the strange habit of breaking off an arm, which then grows a cluster of four new arms that are temporarily in a “comet” form. Sometimes the animal breaks across the central disc; regeneration may result in six‑ or seven‑rayed animals. These divisions seem to be a method of reproduction practiced by the young, for adult animals cease to fragment and produce eggs.
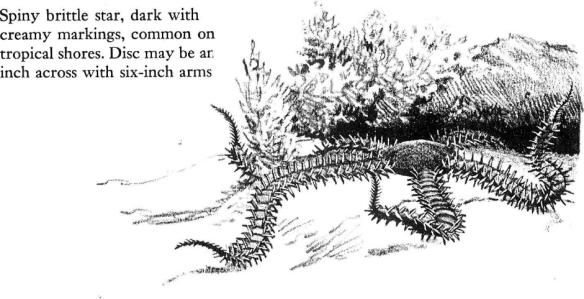
About the bases of gorgonians, under and inside of sponges, under movable rocks and down in little, eroded caverns in the coral rock live the brittle stars. With their long and flexible arms, each composed of a series of “vertebrae” shaped like hourglasses, they are capable of sinuous and graceful motion. Sometimes they stand on the tips of two arms and sway in the motion of the water currents, bending the other arms in movements as graceful as those of a ballet dancer. They creep over the substratum by throwing two of their arms forward and pulling up the body or disc and the remaining arms. The brittle stars feed on minute mollusks and worms and other small animals. In turn, they are eaten by many fish and other predators, and sometimes fall victims to certain parasites. A small green alga may live in the skin of the brittle star; there it dissolves the calcareous plates, so that the arms may break apart. Or a curious little degenerate copepod may live as a parasite within the gonads, destroying them and rendering the animal sterile.
My first meeting with a live West Indian basket star was something I shall never forget. I was wading off Ohio Key in water little more than knee deep when I found it among some seaweeds, gently drifting on the tide. Its upper surface was the color of a young fawn, with lighter shades beneath. The searching, exploring, testing branchlets at the tips of the arms reminded me of the delicate tendrils by which a growing vine seeks out places to which it may attach itself. For many minutes I stood beside it, lost to all but its extraordinary and somehow fragile beauty. I had no wish to “collect” it; to disturb such a being would have seemed a desecration. Finally the rising tide and the need to visit other parts of the flat before they became too deeply flooded drove me on, and when I returned the basket star had disappeared.
The basket starfish or basket fish is related to the brittle stars and serpent stars but displays remarkable differences of structure: each of the five arms diverges into branching V’s, which branch again, and then again and again until a maze of curling tendrils forms the periphery of the animal. Indulging their taste for the dramatic, early naturalists named the basket stars for those monsters of Greek mythology, the Gorgons, who wore snakes in place of hair and whose hideous aspect was supposed to turn men to stone; so the family comprising these bizarre echinoderms is known as the Gorgonocephalidae. To some imaginations their appearance may be “snaky‑locked,” but the effect is one of beauty, grace, and elegance.

All the way from the Arctic to the West Indies basket stars of one species or another live in coastal waters, and many go down to lightless sea bottoms nearly a mile beneath the surface. They may walk about over the ocean floor, moving delicately on the tips of their arms. As Alexander Agassiz long ago described it, the animal stands “as it were on tiptoe, so that the ramifications of the arms form a kind of trellis‑work all around it, reaching to the ground, while the disk forms a roof.” Or again they may cling to gorgonians or other fixed sea growths and reach out into the water. The branching arms serve as a finemeshed net to ensnare small sea creatures. On some grounds the basket stars are not only abundant but associate in herds of many individuals as though for a common purpose. Then the arms of neighboring animals become entwined in a continuous living net to capture all the small fry of the sea who venture, or are helplessly carried, within reach of the millions of grasping tendrils.

To see a basket starfish close inshore is one of those rare happenings that lives always in memory, but it is far otherwise with certain other members of the spiny‑skinned tribe of echinoderms–the holothurians, or sea cucumbers. I have never waded far out onto the flats without meeting them. Their large dark forms, shaped much like the vegetable whose name they have been given, stand out clearly against the white sand where they lie sluggishly, sometimes partly buried. The holothurians perform a function in the sea that is roughly comparable to that of earthworms on land, ingesting quantities of sand and mud and passing it through their bodies. Most of them use a crown of blunt tentacles operated by strong muscles to shovel the bottom sediments into their mouths, then extract food particles from this detritus as it passes through their bodies. Perhaps some calcareous materials are dissolved out by the chemistry of the holothurian body.

Because of their abundance and the nature of their activities, the sea cucumbers profoundly influence the distribution of the bottom deposits around the coral reefs and islands. In a single year, it has been estimated, the holothurians in an area less than two miles square may redistribute 1000 tons of bottom substance. And there is evidence also concerning their work on sea bottoms lying at abyssal depths. The carpeting sediments, which accumulate slowly but unceasingly, lie in orderly layers from which geologists can read many chapters of the past history of the earth. But sometimes the layers are curiously disturbed. Bits of volcanic ash shard originating, for example, from some ancient eruptions of Vesuvius, may in some places lie, not in a thin layer representing and dating the eruption, but widely scattered through the overlying layers of other sediments. Geologists regard this as the work of deep‑sea holothurians. And other evidence from deep dredgings and bottom samplings suggests the existence of herds of holothurians on the sea floor at great depths, working over a bottom area, then moving or in a vast migration directed, not by seasonal change, but by the scarcity of food in those deep and lightless regions.
Except in those parts of the world where they are sought as human food (they are the “trepang,” or beche‑de‑mer, of Oriental markets) the sea cucumbers have few known enemies, yet they possess a strange defense mechanism that they employ when strongly disturbed. Then the holothurian may contract strongly and hurl out the greater part of its internal organs through a rupture in the body wall. Sometimes this action is suicidal, but often the creature continues to live and grows a new set of organs.
Dr. Ross Nigrelli and his associates of the New York Zoological Society have recently discovered that the large West Indian sea cucumber (also found about the Florida Keys) produces one of the most powerful of all known animal poisons, presumably as a chemical means of defense. Laboratory experiments showed that even small doses of the poison affect all kinds of animals, from protozoa to mammals. Fish confined in a tank with the cucumber always die when the act of evisceration occurs. The study of this natural toxin reveals the hazardous existence of many small creatures that live in association with another. The sea
Дата добавления: 2015-05-13; просмотров: 1507;