Гамма-разнообразие наземных экосистем
Уровень – гамма-разнообразие – относится к более крупным пространственным единицам типа острова или ландшафта по сравнению с бета-разнообразием. Затем, если гамма-разнообразие определяется как общее разнообразие группы участков, то эпсилон-разнообразие, или региональное разнообразие, – общее разнообразие группы территорий гамма-разнообразия, которое относится к крупным биогеографическим областям. Наибольший интерес для изучения фитоценохор представляет именно гамма-разнообразие, относящееся к микро-, мезо- и макрокомбинациям растительного покрова, соответствующим урочищам, местностям и ландшафтам в масштабах конкретных геоботанических карт [Виноградов, 2000; Goetze, 2000].
Существующие в литературе характеристики гамма-разнообразия мало способствуют его пониманию. В основополагающем документе [Global Biodiversity Strategy, 1992] отмечено только, что «экосистемное разнообразие (т. е. гамма-разнообразие) труднее измерить, чем видовое или генетическое, поскольку границы сообществ и экосистем менее дискретны». Тем не менее, как только постоянный набор критериев используется, чтобы отличить сообщества и экосистемы, их число и распространение могут быть измерены. Б. А. Юрцев [1992] более определенно различает биохорологическое разнообразие как разнообразие сочетаний организмов тех или иных территориальных выделов, частей биосферы и выделяет биохорологические уровни в пределах экотопа, что соответствует бета-разнообразию ландшафта – гамма-разнообразию биогеографического района – эпсилон-разнообразию. B. C. Безель и др. [1993] определяют, что «исторически сложившееся экосистемное разнообразие какого-либо региона, безусловно, способствует стабилизации круговорота веществ и энергии; появление новых ландшафтов резко разрушает их. В результате такие «новые» экосистемы сами выступают в качестве потребителя ресурсов, резко обедняя их. В итоге биоразнообразие сокращается на всех уровнях организации, включая экосистемный».
По определению Б. В. Виноградова, гамма-разнообразие характеризует информационную диверсификацию фитоценохор на надбиоценотических ландшафтных уровнях. Ввиду пространственной неоднородности и многокомпонентности сложных экосистем, наиболее показательной их характеристикой является пространственно-временное распределение биологических, геофизических, структурных и функциональных свойств, привязанных к содержательным определителям экосистем.
Согласно Шультру и Риклефсу [1993], гамма-разнообразие может быть рассчитано следующим образом:
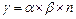 ,
,
где п – общее число местообитаний или ключевых участков, где a и b – среднее значение на ландшафтную единицу. Очевидно, в этом уравнении g -разнообразие может увеличиваться, когда a- и b-разнообразия могут остаться постоянными [Halffler, 1998].
Таким образом, Б. Н. Виноградов рассматривает объектом гамма-разнообразия фитоценохоры, которые, по определению академика В. Б. Сочавы [1972] являются сложными надбиоценотическими пространственными единицами растительного покрова от простых комбинаций растительности урочищ в масштабах 1:30 000 – 1:100 000 до комбинаций растительности ландшафтов в масштабах 1:1 000 000 – 1:3 000 000.
Основным способом исследования гамма-разнообразия фитоценохор является сочетание стационарных и полустационарных описаний пространственных единиц растительного покрова и дешифрирование их пространственно-временных характеристик по аэрокосмическим снимкам. Имеются небольшие опыты такого подхода, немного проясняющие содержание исследования гамма-разнообразия. F. Burel [1993] связывает гамма-разнообразие с фрагментацией экосистем на ландшафтном уровне из-за урбанизации, сельскохозяйственного освоения, вырубки лесов, распознаваемых на космических снимках с SPOT. D. M. Stoms и J. E. Etes [1993] пытаются связать гамма-разнообразие при дистанционных исследованиях с видовым богатством. Причем первый подход основан на последовательной генерализации объекта расчета разнообразия, а второй – на последовательном изменении уровня обработки данных.
Применяются различные показатели для расчета биоразнообразия растительности в пределах ландшафта [Василевич, 1992]. Используются различные уравнения для оценки разнообразия, но наиболее адекватными являются нелинейные (логарифмические) функции, повышающие значение мало распространенных компонент и, наоборот, снижающие подавляющее значение широко распространенных (фоновых) и позволяющие вводить условные определители [Виноградов, 2000].
Привычным для расчета разнообразия является индекс Шеннона, основанный на вычислении простой энтропии.
Для изучения разнообразия фитоценохор считается эффективным использование для исследования гамма-разнообразия вычисления сложной энтропии:
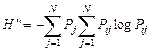 ,
,
где Pj – априорная вероятность появления j-го класса; Рij – вероятность отнесения образца i-го класса к j-му классу.
При многостороннем изучении сложных единиц рекомендуется вычисление условной энтропии Нп [Яглом, 1972] вида:
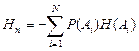 ,
,
где Р(Аi) условные вероятности появления i-го класса по j-й пространственной (или другой условной) характеристике:
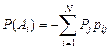 ,
,
а Н(Аi) – частные условные энтропии для каждого класса:
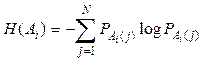 .
.
Применение указанного аппарата в разных направлениях может описать разные свойства гамма-разнообразия. При этом нужно помнить, что все эти показатели не имеют абсолютного значения и используется лишь их сравнение, которое дает прирост информации (DIi), в зависимости от изменения условий опыта, и определяется по разности промежуточных энтропий:
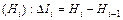 .
.
Во многих работах на ландшафтном уровне биоразнообразие рассматривается как разнообразие сообществ и экосистем в терминах пространственных показателей, включая такие, как степень фрагментации, форма пятен, сложность границ, совмещенность пятен и другие показатели, связанные с измерением ландшафтной структуры [O’Nail et al., 1988, Turner 1990, LaGro, 1991]. Однако такие показатели пространственной неоднородности строго не описаны в информационных терминах.
Дата добавления: 2015-04-11; просмотров: 2413;