Взаимодействие генов
Как правило, взаимодействие генов имеет биохимическую природу, то есть оно основано на взаимодействии белков, синтезируемых под действием определенных генов. Взаимодействовать друг с другом могут как аллельные, так и неаллельные гены.
Взаимодействие аллельных генов. Различают несколько типов взаимодействия аллельных генов: полное доминирование, при котором рецессивный признак не проявляется, неполное доминирование, при котором у гибридов наблюдается промежуточный характер наследования, кодоминирование, в этом случае у гибридов фенотипически проявляются оба признака. Например, кодоминирование проявляется у людей с 4 группой крови. Первая группа крови у людей с аллелями i0i0, вторая — с аллелями IAIA или IAí0; третья — IВIВ или IВí0; четвертая группа имеет аллели IАIВ. Сверхдоминирование – лучшая приспособленность гетерозигот от моногибридного скрещивания (например, Аа) по сравнению с обоими типами гомозигот (АА и аа). Сверхдоминирование можно определить также как гетерозис, возникающий при моногибридном скрещивании. Наиболее известный пример взаимоотношения между нормальным (S) и мутантным (s) аллелями гена, контролирующего структуру гемоглобина у человека. Люди, гомозиготные по мутантной аллели (ss), страдают тяжёлым заболеванием крови – серповидноклеточной анемией, от которого они гибнут обычно в детском возрасте (эритроциты больного имеют серповидную форму и содержат гемоглобин, структура которого незначительно изменена в результате мутации). Однако в тропической Африке и других районах, где распространена малярия, в популяциях человека постоянно присутствуют все три генотипа SS, Ss и ss (20-40% населения гетерозиготы Ss). Оказалось, что сохранение в популяциях человека летальной (смертельной) аллели (s) обусловлено тем, что гетерозиготы (Ss) более устойчивы к малярии, чем гомозиготы по нормальному гену (SS), и, следовательно, обладают отборным преимуществом.
|
 Рис. 331. Наследование формы гребня у кур.
Рис. 331. Наследование формы гребня у кур.
Комплементарное взаимодействие. Комплементарными называют гены, обусловливающие при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новое фенотипическое проявление признака. Классическим примером комплементарного взаимодействия генов является наследование формы гребня у кур (рис. 331). При скрещивании кур, имеющих розовидный и гороховидный гребень, все первое поколение имеет ореховидный гребень.
При скрещивании гибридов первого поколения у потомков наблюдается расщепление по форме гребня: 9 ореховидных: 3 розовидных: 3 гороховидных: 1 листовидный. Генетический анализ показал, что куры с розовидным гребнем имеют генотип А_bb, с гороховидным — ааВ_, с ореховидным — А_В_ и с листовидным — ааbb, то есть развитие розовидного гребня происходит в том случае, если в генотипе имеется только один доминантный ген — А, гороховидного — наличие только гена В, сочетание генов А В обусловливает появление ореховидного гребня, а сочетание рецессивных аллелей этих генов — листовидного.
При комплементарном взаимодействии генов в дигибридном скрещивании получаются расщепления потомков отличные от менделевского: 9:7, 9:3:4, 13:3, 12:3:1, 15:1, 10:3:3, 9:6:1. Однако все они являются видоизменениями общей менделевской формулы 9:3:3:1.
|
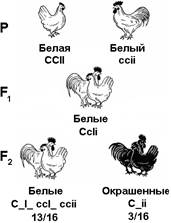 Рис. 332. Наследование окраски оперения у кур
Рис. 332. Наследование окраски оперения у кур
Белое оперение определяется несколькими различными генами, например, у белых леггорнов — генами ССII, а у белых плимутроков — ccii (рис. 332). Доминантная аллель гена С определяет синтез предшественника пигмента (хромогена, обеспечивающего окраску пера), а его рецессивная аллель с — отсутствие хромогена. Ген I является подавителем действия гена С, а аллель i не подавляет его действия. Таким образом, белая окраска у кур определяется не наличием особых генов, определяющих развитие этой окраски, а наличием гена, подавляющего ее развитие.
При скрещивании, например, леггорнов (ССII) с плимутроками (ссii), все потомство F1 имеет белую окраску, которая определяется наличием в их генотипе гена-подавителя (СсIi). Если же гибридов F1 скрестить между собой, то во втором поколении происходит расщепление по окраске в отношении 13/16 белых: 3/16 окрашенных. Окрашенным оказывается та часть потомства, в генотипе которой имеется ген окраски и отсутствует его подавитель (С_ii).
Полимерия. Скрещивая белую и пурпурную фасоли, Мендель столкнулся с явлением полимерии. Полимерией называют однозначное влияние двух, трех и более неаллельных генов на развитие одного и того же признака. Такие гены называют полимерными, или множественными, и обозначают одной буквой с соответствующим индексом, например, А1, А2, а1, а2. Полимерные гены контролируют большинство оличественных признаков организмов: высоту растения, массу семян, масличность семян, содержание сахара в корнеплодах сахарной свеклы, удойность коров, яйценоскость, вес тела и т.д.
Явление полимерии было открыто в 1908 г. при изучении окраски зерновки у пшеницы Нельсоном-Эле (рис. 333). Он предположил, что наследование окраски у зерновки пшеницы обусловлено двумя или тремя парами полимерных генов. При скрещивании краснозерной и белозерной пшеницы в F1 наблюдалось промежуточное наследование признака: все гибриды первого поколения имели светло-красное зерно. В F2 происходило расщепление в отноше
|
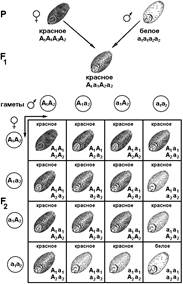 Рис. 333. Наследование окраски зерновки у пшеницы.
Рис. 333. Наследование окраски зерновки у пшеницы.
Причем краснозерные зерновки имели разную интенсивность окраски — от темно-красной до светло-красной. Исходя из наблюдений, Нельсоном-Эле определил, что признак окраски зерновок обуславливает три пары полимерных генов. У человека по типу полимерии наследуется, например, окраска кожи.
Плейотропия. Плейотропией называют множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака, но и воздействует на вторичные реакции биосинтеза различных других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который обнаружил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь—серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают глубокие нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию погибает в детстве.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
Дата добавления: 2015-04-10; просмотров: 2241;