ПОТЕНЦИАЛ ПОКОЯ
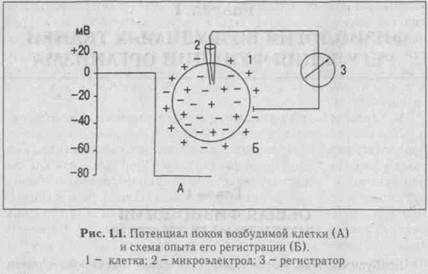
А. Общая характеристика и непосредственная причина формирования. Потенциал покоя (ПП) - относительно стабильная разность между величинами электрических потенциалов внутри и вне клетки в покое. Его величина обычно варьирует в пределах 30-90 мВ (в волокнах скелетной мышцы - 60-90 мВ, в нервных клетках -50-80 мВ, в гладких мышцах -30-70 мВ, в сердечной мышце -80-90 мВ). При регистрации ПП луч осциллографа во время прокола мембраны клетки микроэлектродом скачком отклоняется и показывает отрицательный заряд внутри клетки (рис. 1.1).
ПП играет исключительно важную роль в жизнедеятельности самой клетки и организма в целом. В частности, он составляет основу для переработки информации нервной клеткой, обеспечи-* вает регуляцию деятельности внутренних органов и опорно-двигательного аппарата посредством запуска процессов возбуждения и сокращения в мышце. Нарушение процессов возбуждения в кардио-миоцитах ведет к остановке сердца. Согласно мембранно-ионной теории (Бернштейн, Ходжкин, Хаксли, Катц, 1902-1952), непосредственной причиной формирования ПП является неодинаковая концентрация анионов и катионов внутри и вне клетки.
В нервных и мышечных клетках концентрация К+ внутри клетки в 30-40 раз больше, чем вне клетки; концентрация №+ вне клетки в 10-12 раз больше, нежели внутри клетки; ионов С1~ вне клетки в 15-20 раз больше, чем внутри клетки. В клетке имеется небольшое количество ионов М§2+. Кальций в свободном состоянии находится в основном вне клетки. Он содержится также в эн-доплазматическом ретикулуме; в гиалоплазме его очень мало. Это обусловливается отчасти активным транспортом Са2+ наружу через клеточную мембрану, отчасти поглощением его эндоплазмати-ческим ретикулумом - это резервуар для Са2+- и другими органел-лами, например, митохондриями, связыванием Са2+ цитратом, глютаматом.
В клетке находятся также крупномолекулярные анионы - главным образом это отрицательно заряженные белковые молекулы,
например, глютамат, аспартат, а также органические фосфаты. Различные ионы распределены неравномерно по обе стороны клеточной мембраны, во-первых, вследствие неодинаковой проницаемости клеточной мембраны для различных ионов, во-вторых, в результате работы ионных насосов, транспортирующих ионы в клетку и из клетки вопреки концентрационному и электрическому градиентам.
Б. Терминология и характеристика клеточной мембраны.
Проницаемость клеточной мембраны - это ее способность пропускать воду, незаряженные и заряженные частицы (ионы) согласно законам диффузии и фильтрации. Проницаемость клеточной мембраны (она представляет собой липопротеиновую оболочку) определяется следующими факторами: 1) наличием в составе мембраны различных ионных каналов - управляемых (с воротным механизмом) и неуправляемых (каналов утечки); 2) размерами каналов и размерами частиц; 3) растворимостью частиц в мембране (клеточная мембрана проницаема для растворимых в ней липи-дов и непроницаема для пептидов).
Проводимость - это способность заряженных частиц (ионов) проходить через клеточную мембрану согласно электрохимическому градиенту. Проводимость иона зависит от его электрохимического градиента и от проницаемости мембраны - чем они больше, тем лучше проводимость иона через мембрану. Перемещения ионов в клетку и из клетки, согласно концентрационному и электрическому градиентам в состоянии покоя клетки, осуществляются преимущественно через неуправляемые (без воротного механизма) каналы, их называют также каналами утечки. Неуправляемые каналы всегда открыты, они практически не меняют своей пропускной способности при электрическом воздействии на клеточную мембрану и ее возбуждении. Неуправляемые каналы подразделяются на ионоселективные каналы (например, калиевые медленные неуправляемые каналы) и иононеселективные каналы. Последние пропускают различные ионы - К+, Ыа+, СГ.
В. Роль проницаемости клеточной мембраны и различных ионов в формировании ПП. №+ и К+ в покоящейся клетке перемещаются через мембрану согласно законам диффузии, при этом К+ из клетки выходит в значительно большем количестве, чем входит Ыа+ в клетку, поскольку проницаемость клеточной мембраны для К+ примерно в 25 раз больше проницаемости для Ыа+. Органические анионы из-за своих больших размеров не могут выходить из клетки, поэтому внутри клетки в состоянии покоя отрицательных ионов оказывается больше, чем положительных. Однако превышение абсолютного числа анионов над числом катио-
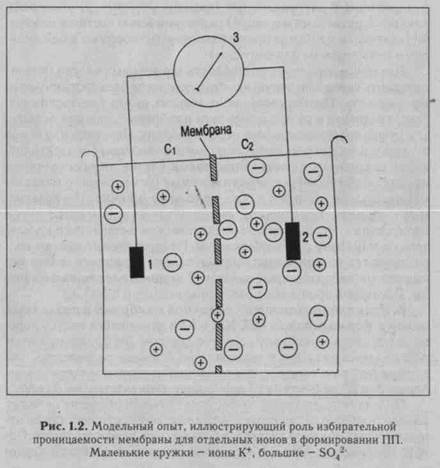
нов в клетке чрезвычайно мало. Но этого различия достаточно для создания разности электрических потенциалов внутри и вне клетки. Главным ионом, обеспечивающим формирование ПП, является ион К+ (рис. 1.2). Об этом свидетельствуют результаты опыта с перфузией внутреннего содержимого гигантского аксона кальмара солевыми растворами. При уменьшении концентрации К+ в перфузате ПП уменьшается, при увеличении концентрации К+ ПП увеличивается. В покоящейся клетке устанавливается динамическое равновесие между числом выходящих из клетки и входящих в клетку ионов К+. Электрический и концентрационный градиенты противодействуют друг другу - согласно концентрационному градиенту К+ стремится выйти из клетки, отрицательный заряд внутри клетки и положительный заряд наружной поверхности клеточ-
ной мембраны препятствуют этому. Когда концентрационный и электрический градиенты уравновесятся, число выходящих из клетки ионов К+ сравняется с числом входящих в клетку ионов К+. В этом случае на клеточной мембране устанавливается так называемый равновесный калиевый потенциал.
Равновесный потенциал для любого иона можно рассчитать по формуле Нернста. Концентрация положительно заряженного иона, находящегося снаружи, в формуле Нернста располагается в числителе, иона, находящегося внутри клетки, - в знаменателе. Для отрицательно заряженных ионов расположение противоположное:
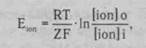
|
где Е(оп - потенциал, создаваемый данным ионом; К - газовая постоянная (8,31 Дм); Т - абсолютная температура (273+37°С); 2 -валентность иона; Р - постоянная Фарадея (9,65 104); [юп]\ - концентрация иона внутри клетки ОпзШе); [юп]о - концентрация иона во внешней среде клетки (оиЫае).
Вклад Л^а+ и СГ в создание ПП. Проницаемость клеточной мембраны в покое для Ыа+ очень низкая - намного ниже, чем для К+, тем не менее она имеет место, поэтому ионы №+, согласно концентрационному и электрическому градиентам, стремятся и в небольшом количестве проходят внутрь клетки. Это ведет к уменьшению ПП, так как на внешней поверхности клеточной мембраны суммарное число положительно заряженных ионов уменьшается, хотя и незначительно, а часть отрицательных ионов внутри клетки нейтрализуется входящими в клетку положительно заряженными ионами №+. Что касается СГ, его влияние на величину ПП противоположно влиянию №+ и зависит от проницаемости клеточной мембраны для СГ (она в 2 раза ниже, чем для К+). Дело в том, что СГ, согласно концентрационному градиенту, стремится и проходит в клетку, что ведет к увеличению ПП. Концентрации ионов К+ и СГ близки между собой. Но СГ находится в основном вне клетки, а К+ - внутри клетки. Препятствует входу СГ в клетку электрический градиент, поскольку заряд внутри клетки отрицательный, как и заряд СГ. Наступает равновесие сил концентрационного градиента, способствующего входу СГ в клетку, и электрического градиента, препятствующего входу СГ в клетку.
Г. Определенную роль в формировании ПП играют поверхностные заряды самой клеточной мембраны и ионы Са2+. Наружная и внутренняя поверхности клеточной мембраны несут собственные электрические заряды, преимущественно с от-
рицательным знаком. Это полярные молекулы клеточной мембраны: гликолипиды, фосфолипиды, гликопротеиды. Фиксированные наружные отрицательные заряды, нейтрализуя положительные заряды внешней поверхности мембраны, уменьшают ПП. Фиксированные внутренние отрицательные заряды клеточной мембраны, напротив, суммируясь с анионами внутри клетки, увеличивают ПП. Роль ионов Са2+ в формировании ПП заключается в том, что они взаимодействуют с наружными отрицательными фиксированными зарядами мембраны клетки и отрицательными карбоксильными группами интерстиция и нейтрализуют их, что ведет к увеличению и стабилизации ПП.
Таким образом, ПП- это алгебраическая суммане только всех зарядов ионов вне и внутри клетки, но также алгебраическая сумма отрицательных внешних и внутренних поверхностных зарядов самой мембраны.
При проведении измерений потенциал окружающей клетку среды принимают за величину, равную нулю. Относительно нулевого потенциала внешней среды потенциал внутренней среды клетки, как отмечалось выше, составляет величину порядка 60-90 мВ. Повреждение клетки приводит к повышению проницаемости клеточных мембран, в результате чего различие проницаемости для,, К+ и Ка+ уменьшается, ПП при этом снижается.
Д. Роль ионных насосов в формировании ПП. В результате непрерывного перемещения различных ионов через клеточную мембрану их концентрация внутри и вне клетки постепенно должна выравниваться. Однако, несмотря на постоянную диффузию ионов (утечку ионов), ПП клеток остается на одном уровне. Следовательно, кроме собственно ионных механизмов формирования ПП, связанных с различной проницаемостью клеточной мембраны, имеется активный механизм поддержания градиентов концентрации различных ионов внутри и вне клетки. Им являются ионные насосы, в частности Ыа/К-насос (помпа).
Ионный насос - это транспортная система, обеспечивающая перенос иона с непосредственной затратой энергии вопреки концентрационному и электрическому градиентам. Если заблокировать освобождение энергии, например динитрофенолом, в течение 1 ч выведение Ыа+ из клетки сократится примерно в 100 раз. Как выяснилось, выведение №+ сопряжено с транспортом К+, что можно продемонстрировать при удалении К+ из наружного раствора. Если К+ на наружной стороне мембраны нет, работа насоса блокируется, перенос Ыа+ из клетки в этом случае падает, составляя примерно 30% от нормального уровня. Сопряженность транспорта Ыа+ и К+ уменьшает расход энергии примерно в 2 раза по сравне-
нию с той, которая потребовалась бы при несопряженном транспорте. В целом, траты энергии на активный транспорт веществ огромны - лишь Ыа /К- насос потребляет 1 / 3 всей энергии, расходуемой организмом в покое. За 1 с один Ыа/К-насос (одна молекула белка) переносит 150-600 ионов №+. Накопление №+ в клетке стимулирует работу №/К-насоса, уменьшение Ыа+ в клетке снижает его активность, поскольку снижается вероятность контакта ионов с соответствующим переносчиком. В результате сопряженного транспорта Ыа+ и К+ поддерживается постоянная разность концентраций этих ионов внутри и вне клетки. Одна молекула АТФ обеспечивает один цикл работы №/К-насо-са - перенос трех ионов Ыа+ за пределы клетки и двух ионов К+ внутрь клетки. Асимметричный перенос ионов №/К- насосом поддерживает избыток положительно заряженных частиц на наружной поверхности клеточной мембраны и отрицательных зарядов внутри клетки, что позволяет считать № / К-насос структурой электрогенной, дополнительно увеличивающей ПП примерно на 5-10 мВ (в среднем около 10% у разных возбудимых клеток -у одних больше, у других меньше). Данный факт свидетельствует о том, что решающим фактором в формировании ПП является селективная проницаемость клеточной мембраны для разных ионов. Если уравнять проницаемость клеточной мембраны для всех ионов, то ПП будет составлять только 5-10 мВ - за счет работы Ыа/К-помпы.
Нормальная величина ПП является необходимым условием возникновения процесса возбуждения клетки, т. е. возникновения и
аспространения потенциала действия, инициирующего специфи-
ескую деятельность клетки.
Дата добавления: 2015-02-23; просмотров: 916;