ЛЕКЦИЯ 5. ОБЩАЯ ФИЗИОЛОГИЯ НЕРВНОЙ СИСТЕМЫ
План
1. Механизм проведения возбуждения.
2. Синаптическая передача нервного импульса.
3. Свойства нервных центров.
4. Координирующая роль ЦНС.
5. Торможение в ЦНС.
Механизм проведения возбуждения. Нервные импульсы распространяются по нервным волокнам от рецепторов к нервному центру или от нервного центра к эффектору. Нервные волокна делят на три класса: А, В и С. К классу А относятся толстые миелиновые волокна, скорость проведения возбуждения в них достигает 70-120 м/c. К классу В относятся миелинизированные преганглионарные волокна вегетативной нервной системы, скорость проведения возбуждения в них 3-14 м/c. Волокна класса С – безмиелиновые постганглионарные волокна вегетативной нервной системы. Скорость проведения возбуждения – 0,5-2 м/с.
Механизм проведения возбуждения в нервных волокнах объясняется возникновением локальных токов, появляющихся между возбужденным и невозбужденным участками мембраны нервного волокна.
В безмиелиновых волокнах возбуждение распространяется непрерывно, а в миелинизированных волокнах – скачками между перехватами Ранвье.
Синаптическая передача нервного импульса.Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). В нервной системе синапсы образуются между отростками разных нейронов, а так же между отростками и телами клеток. Соответственно их называют аксо-аксональными, аксо-дендритными, аксо-соматическими, дендро-дендритными. Передача возбуждения с отростка одной нервной клетки на отросток или тело другой нервной клетки возможна двумя способами: электрическим и химическим. Возбуждающим электрическим синапсам свойственна очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран для прохождения через них электрического тока. Такие синапсы находятся в сердечной мышце, гладких мышцах и железистой ткани. Основным способом передачи возбуждения между нервными клетками является химический. Синапс с химической передачей состоит из синаптической терминали, пресинаптической мембраны, синаптической щели и постсинаптической мембраны.
В пресинаптическом окончании синапса находятся митохондрии и пресинаптические пузырьки (визикулы) c медиатором. Синаптическая щель имеет ширину 20-50 нм. Постсинаптическая мембрана утолщена и образована мембраной иннервируемой клетки, а в концевой пластинке – мембраной мышечного волокна.
Механизм синаптической передачи.Под влиянием нервного импульса наступает деполяризация окончаний аксона, что повышает в нем концентрацию Ca 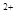 , и содержание синаптических пузырьков выбрасывается в синаптическую щель. Медиатор (ацетилхолин) диффундирует в синаптическую щель и связывается с рецепторами постсинаптической мембраны. Действие молекул медиатора ведет к открытию ионных каналов для Na
, и содержание синаптических пузырьков выбрасывается в синаптическую щель. Медиатор (ацетилхолин) диффундирует в синаптическую щель и связывается с рецепторами постсинаптической мембраны. Действие молекул медиатора ведет к открытию ионных каналов для Na 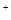 и К , что вызывает возникновение возбуждающего постсинаптического потенциала (ВПСП). В тормозных синапсах, тормозный медиатор (гамма-аминомасляная кислота) открывает в постсинаптической мембране селективные хлорные каналы. При этом возникает тормозный постсинаптический потенциал (ТПСП) .
и К , что вызывает возникновение возбуждающего постсинаптического потенциала (ВПСП). В тормозных синапсах, тормозный медиатор (гамма-аминомасляная кислота) открывает в постсинаптической мембране селективные хлорные каналы. При этом возникает тормозный постсинаптический потенциал (ТПСП) .
Свойства нервных центров.Группа нейронов, необходимых для осуществления определенного рефлекса или управления одной из функций организма, образует нервный центр. Свойства нервных центров зависят от строения и механизма передачи возбуждения в синапсах: 1.Одностороннее проведение возбуждения. В нервном центре, в отличии от нервного волокна, возбуждение распространяется только в одном направлении: от центростремительного нейрона к центробежному. 2. Замедление проведение возбуждения. Это свойство вызвано тем, что в нервном центре может быть несколько нейронов и, следовательно, столько же синапсов. В каждом синапсе происходит синаптическая задержка проведения возбуждения. 3. Суммация. Заключается в накоплении (сложении) эффектов подпороговых раздражений. Одно пдпороговое раздражение не вызывает ответной рефлекторной реакции, а несколько подпороговых раздражений в сумме дают нужный эффект. 4. Трансформация ритма возбуждения. Нервные центры обладают способностью трансформировать частоту и ритм поступающих импульсов. На одиночное раздражение, поступившее в нервный центр, он может ответить серией импульсов. 5. Последействие. Ответная рефлекторная реакция продолжается некоторое время после прекращения действия раздражителя. Причиной последействия являются следовая деполяризация и циркуляция нервных импульсов по кольцевым связям между нейронами данного центра. 6. Утомляемость нервного центра. Утомление в ЦНС возникает в нервном центре. Это обусловлено его низкой лабильностью. Утомление в нервном центре проявляется в постепенном уменьшении, а затем и прекращении рефлекторного ответа при действии раздражителя. Причиной возникновения утомления является нарушение передачи возбуждения в синапсах. 7. Пластичностьнервных центров. В определенных условиях нервные центры перестраиваются и приобретают новые, несвойственные им ранее функции. Это особенно ярко проявляется при операциях удаления различных отделов мозга.
Координирующая роль ЦНС. Приспособление организма к различным изменениям внешней среды возможно благодаря наличию в ЦНС координации функций. Под координацией функций понимают взаимодействие нейронов, а следовательно и нервных процессовв ЦНС, которое обеспечивает ее согласованную деятельность, направленную на интеграцию (объединение) функций различных органов и систем организма. Известен ряд механизмов, лежащих в основе координирующей деятельности нервной системы. Одни из них связаны с морфологическими особенностями ее строения (принцип общего конечного пути, принцип обратных связей), другие – с функциональными свойствами (иррадиация, индукция, доминанта).
Принцип общего конечного пути. Был открыт Ч.С. Шеррингтоном. Этот принцип исходит из анатомического соотношения между афферентными и эфферентными нейронами. Количество чувствительных нейронов, приносящих возбуждение в ЦНС в 5 раз больше, чем двигательных. Поэтому, к одному мотонейрону поступают импульсы от многих рецепторов, расположенных в различных частях тела. Этот процесс называется конвергенцией. Таким образом, самые разнообразные стимулы могут быть причиной одной и той же рефлекторной реакции, т.е. происходит борьба за “общий конечный путь”.
Принцип обратных связей. Воздействие работающего органа на состояние управляющего им центра называется обратной связью. Различают положительные и отрицательные обратные связи. Если импульсы, возникающие в результате какой-либо рефлекторной реакции, поступая в управляющий ею нервный центр, усиливают ее, - это положительная обратная связь. Если они угнетают эту реакцию, то это отрицательная обратная связь.
Иррадиация. Возбуждение, возникшее в одном из нервных центров, способно распространятся по ЦНС, возбуждая новые участки. Процесс распространения возбуждения называют иррадиацией. Она обусловлена наличием многочисленных связей между нейронами в ЦНС.
Индукция. Процессы возбуждения и торможения в ЦНС находятся в определенных отношениях, которые осуществляются по законам индукции. Возбуждение, возникшее в одном нервном центре, “наводит” торможение на другой, и наоборот.
Принципдоминанты. Был разработан А.А. Ухтомским. Господствующий очаг возбуждения, определяющий характер ответных реакций организма на внешние и внутренние раздражения, Ухтомский назвал доминантой. Доминантный очаг возбуждения характеризуется признаками: 1) повышенной возбудимостью; 2) стойкостью возбуждения; 3) повышенной способностью к суммации возбуждения; 4) инерцией, т.е. способностью длительно сохранять возбуждение после окончания действия раздражителя; 5) способностью вызывать сопряженное торможение.
Торможение в ЦНС. Торможение – это особый нервный процесс проявляющийся в уменьшении или полном исчезновении ответной реакции. Процесс торможения в ЦНС был открыт И.М. Сеченовым (1862). В ЦНС наряду с возбуждающими имеются и тормозные нейроны. На каждой нервной клетке располагаются возбуждающие и тормозные синапсы. Поэтому, в каждый момент на теле нейрона в одних синапсах возникает возбуждение, а в других – торможение.
По характеру возникновения различают первичное и вторичное торможение.
Первичное торможение возникает под влиянием раздражения сразу без предварительного возбуждения и осуществляется с участием тормозных синапсов. Вторичное торможение осуществляется без участия тормозных структур и возникает вследствие перехода возбуждения в торможение. Первичное торможение по месту возникновения может быть пресинаптическим и постсинаптическим. Пресинаптическое торможение осуществляется перед прохождением импульса через синапс. Постсинаптическое торможение может быть прямым и возвратным. Прямое торможение осуществляется вставочными нейронами спинного мозга, корзинчатыми нейронами таламуса, тормозными клетками мозжечка, а возвратное торможение через специальные тормозные нейроны – клетки Реншоу.
Дата добавления: 2015-02-05; просмотров: 2366;