ЕВРАЗИЙСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ ИМЕНИ Л.Н.ГУМИЛЕВА
Ассимиляты из листьев поступают во флоэму, состоящую из нескольких типов клеток, специализированных в метаболическом и структурном отношениях. Транспортную функцию осуществляют ситовидные клетки (у низших сосудистых растений и голосеменных) или ситовидные трубки (у покрытосеменных растений). Паренхимные клетки-спутники выполняют энергетическую функцию; другие паренхимные клетки пучка являются передаточными или запасающими. Все эти типы клеток функционируют во взаимодействии.
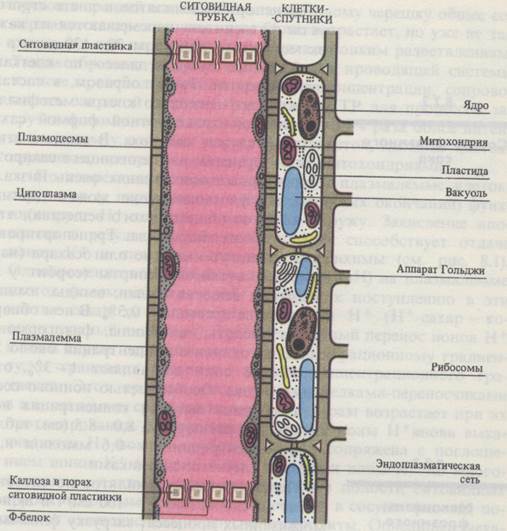
Ситовидные элементы. В ситовидных трубках плазмалемма окружает протопласт, содержащий небольшое число митохондрий и пластид, а также агранулярный эндоплазматический ретикулум. Тонопласт в ситовидных элементах разрушается. Зрелая ситовидная трубка лишена ядра. Поперечные клеточные стенки - ситовидные пластинки - имеют перфорации, выстланные плазмалеммой и заполненные полисахаридом каллозой и фибриллами актиноподобного Ф-белка, которые ориентированы продольно. Ситовидные трубки многочисленными плазмодесмами связаны с клетками-спутниками.
Сопровождающие клетки. Клетки-спутники, или сопровождающие клетки, - это небольшие вытянутые вдоль ситовидных трубок паренхимные клетки с крупным ядром, электронноплотной цитоплазмой, с большим количеством рибосом, других органелл и особенно митохондрий, обилие которых создает структуру, называемую митохондриальным ретикулумом. Клеточные стенки этих клеток пронизаны ветвящимися плазмодесмами, собранными в утолщениях стенок (плазмодесменных полях). Число плазмодесм в сопровождающих клетках в 3- 10 раз больше, чем в стенках соседних клеток мезофилла листа (например, в листе ясеня общее число плазмодесм на клетку мезофилла 800 - 3000, а в клетке-спутнике 24000). Другая особенность клеточных стенок клеток-спутников - обилие в них инвагинаций («лабиринтов»), выстланных плазмалеммой, что значительно увеличивает ее поверхность.
Передвижение ассимилятов по ситовидным трубкам происходит со скоростью 50-100 см/ч и включает в себя три взаимосвязанных процесса: загрузку флоэмы, транспорт ассимилятов по ситовидным элементам и разгрузку флоэмы.
Состав флоэмного сока. Основной транспортной формой сахаров у большинства растений служит сахароза. В связи с этим активность инвертазы - фермента, расщепляющего сахарозу на глюкозу и фруктозу, в проводящих тканях очень низка. Концентрация сахарозы во флоэмном соке может достигать 0,8-1,0 моль/л (80 - 85% от общего сухого вещества), что в 10 - 30 раз выше, чем в клетках мезофилла. Транспортироваться по ситовидным трубкам могут и другие олигосахара (например, у ясеня - рафиноза и стахиоза), спирты (сорбит у яблони). Содержание азотистых веществ (белки, амиды, аминокислоты) во флоэмном соке не превышает 0,5%. В нем обнаружены также органические кислоты, витамины, фитогормоны (ауксин, гиббереллины, цитокинин в концентрации около 5 - 50 нмоль/л). Неорганические соли составляют 1-— 3 % от общего количества веществ сока. Особенностью ионного состава флоэмного эксудата является высокая концентрация ионов К+ (до 50 - 200 ммоль/л) и рН порядка 8,0 - 8,5. АТР содержится в концентрации 0,4 - 0,6 ммоль/л и, по-видимому, поставляется клетками-спутниками.
Загрузка флоэмных окончаний. В свободном пространстве клеточных стенок (в апопласте) может находиться до 20% сахаров, содержащихся в листе, куда они попадают главным образом в виде сахарозы из фотосинтезирующих клеток мезофилла. Особенности транспорта ассимилятов от мезофилла к флоэме изучены недостаточно, но, по-видимому, у разных видов растений он происходит неодинаково. Если в стенках клеток обкладки есть пояски Каспари, то сахара в этих участках должны проходить через симпласт. Многочисленные плазмодесмы между клетками обкладки или клетками листовой паренхимы и клетками-спутниками могут способствовать симпластному транспорту. Развитие системы «лабиринтов» в стенках клеток, лежащих между мезофиллом и ситовидными трубками (у ряда представителей бобовых), должно способствовать транспорту ассимилятов через мембрану и апопластному перемещению веществ. У некоторых растений (сахарная свекла) нет структурных приспособлений для облегчения передвижения ассимилятов. В этом случае транспорт может быть обеспечен сильно развитой системой переносчиков на мембранах клеток.
Измерение осмотической концентрации сахаров показало, что в клетках мезофилла осмотическое давление ниже, чем в тонких проводящих пучках. По мере продвижения от тонких пучков к средней жилке и далее к листовому черешку общее содержание сахаров в пучках еще более возрастает, но уже не так резко, как при переходе от мезофилла к тонким разветвлениям. Таким образом, загрузка ассимилятами проводящей системы в листе происходит против градиента концентрации, сопровождаясь затратой энергии. Источником АТР для процессов загрузки флоэмы, по-видимому, служит в 3 - 4 раза более интенсивное, чем у соседних паренхимных клеток, дыхание клеток-спутников, которые особенно богаты митохондриями.
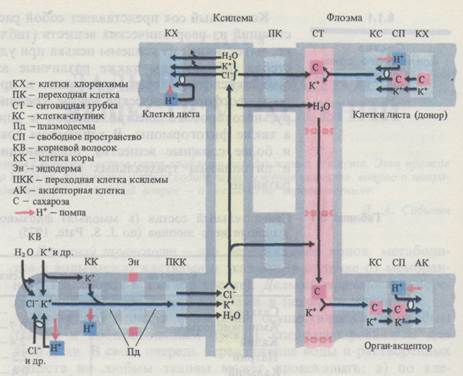
По современным представлениям, в плазмалемме клеток-спутников (у паренхимных клеток флоэмных окончаний) функционирует Н+-помпа, направленная наружу. Закисление апопласта в результате работы Н+-помпы способствует отдаче ионов К+ и сахарозы клетками хлоренхимы. Одновременно возникший градиент рН (ΔрН) на плазмалемме клеток флоэмных окончаний приводит к поступлению в эти клетки сахарозы в симпорте с ионами Н+ (Н+-сахар - котранспортный механизм). Трансмембранный перенос ионов Н+ в этом случае осуществляется по концентрационному градиенту, а транспорт сахарозы - против концентрационного градиента. Этот процесс обеспечивается белками-переносчиками в плазмалемме, сродство которых к сахарам возрастает при их протонировании. Поступившие в клетки ионы Н+ вновь выкачиваются Н+-помпой, работа которой сопряжена с поглощением ионов К+). Сахароза и ионы К+ по многочисленным плазмодесмам переносятся в полости ситовидных трубок. Сходным образом из мезофилла в сосуды флоэмы попадают аминокислоты и другие метаболиты. Описанный механизм объясняет не только накопление сахарозы в ситовидных элементах, но и то обстоятельство, что в ситовидных трубках поддерживаются высокое содержание калия и низкая концентрация протонов. Возрастание концентрации осмотически активных веществ приводит к притоку в полость ситовидных трубок воды из окружающих тканей, прежде всего из сосудов ксилемы.
Флоэмный сок содержит разнообразные ионы. С помощью радиоактивных изотопов установлено, что из ксилемного сока во флоэмный легко циркулируют калий, натрий, магний, фосфор, азот. Микроэлементы - железо, марганец, цинк, молибден - способны перемещаться по флоэме из зрелых в молодые растущие листья. Циркуляция ионов между ксилемой и флоэмой осуществляется следующим образом. В ситовидную трубку ионы могут попадать как путем транспорта из клеток мезофилла, так и не выходя за пределы листовой жилки, при участии паренхимных клеток, в клеточных стенках которых образуются многочисленные выросты цитоплазмы («лабиринты»). Клетки с таким строением стенки, как правило, активно участвуют в транспорте ионов и называются переходными (передаточными). «Лабиринты» в стенках могут быть поляризованы. Например, у переходных клеток ксилемы стебля и листовых черешков выросты цитоплазмы есть лишь в стенке, примыкающей к сосуду, что может обеспечивать перенос ионов из ксилемного сока в симпласт ксилемной паренхимы. В переходных клетках, расположенных в жилке между сосудом ксилемы и ситовидной трубкой, выросты имеются на стенках, смежных с обоими типами транспортных систем. Сходный обмен ионами (и водой) может осуществляться и в стебле, где проводящие элементы ксилемы и флоэмы также сближены.
Транспорт веществ по ситовидным трубкам. Передвижение ассимилятов по ситовидным элементам - особый случай симпластического транспорта. Механизм этого транспорта до конца не раскрыт.
Наиболее обоснована предложенная Э. Мюнхом (1926) гипотеза потока под давлением. Согласно этой гипотезе между фотосинтезирующими клетками листа, где в симпласте накапливается сахароза, и тканями, использующими ассимиляты (например, корнем), создается осмотический градиент, который в ситовидных трубках превращается в градиент гидростатического давления. В результате во флоэме возникает ток жидкости под давлением от листа к корню. В настоящее время эта гипотеза получила поддержку благодаря изучению механизмов загрузки и разгрузки флоэмы - решающих факторов дальнего транспорта ассимилятов. Сохраняющаяся в ситовидных трубках плазмалемма с ее свойством избирательной проницаемости является важнейшим условием, поддерживающим поток под давлением. В какой-то мере описанный механизм флоэм-ного транспорта сходен с механизмом, обеспечивающим подъем пасоки по ксилеме под действием корневого давления. Сходство особенно очевидно в случае весеннего «плача» древесных, в пасоке которых в этот период содержится много сахаров.
Возможно, однако, что транспорт по флоэме на дальние расстояния обеспечивается не единственным механизмом. Предполагается, что движущей силой перемещения флоэмной жидкости из одной ситовидной трубки в другую через поры может быть транспорт ионов К+ в ходе электроосмотического процесса. Согласно этой гипотезе, К+ активно входит в ситовидную трубку выше ситовидной пластинки и выходит в апопласт в следующем членике ниже ситовидной пластинки. В результате на ситовидных пластинках возникает отрицательный электрический градиент, способствующий транспорту через них ионов К+, а с ними и молекул воды.
Предполагается также, что фибриллы актиноподобного Ф-белка в порах ситовидных пластинок во взаимодействии с миозином могут обладать контрактильными (сократительными) свойствами, что способствует передвижению жидкости по флоэме. Однако обе эти гипотезы - электроосмотическая и контрактильная - не имеют пока убедительных экспериментальных доказательств.
Разгрузка флоэмы. Для разгрузки флоэмы существенное значение имеет повышенное гидростатическое давление в ситовидных трубках. Помимо этого интенсивно растущие и запасающие органы обладают аттрагирующей силой, т.е. способны поглощать ассимиляты из флоэмы. В плазмалемме клеток акцепторной зоны функционирует Н+-помпа, которая, с одной стороны, воздействует на ситовидные трубки и клетки-спутники, закисляя апопласт и таким образом побуждая их отдавать ионы К+ и сахарозу в фазу клеточных стенок, а с другой - создает условия для поступления сахаров и калия из апопласта в симпласт акцепторных тканей. Сахароза поглощается с участием мембранных переносчиков в симпорте с протонами, а ионы К+ - по электрическому градиенту. В клетках запасающих органов, например в корнеплоде сахарной свеклы, сахароза накапливается в вакуолях. Предполагается, что через тонопласт активно переносятся ионы К+, а это способствует транспорту сахарозы. Конкретные механизмы поступления К+ и сахарозы в вакуоль неизвестны.
Регуляция флоэмного транспорта. Транспорт ассимилятов из цитоплазмы клеток мезофилла листа (донор) к клеткам органов-акцепторов, например корня, включает в себя ряд транспортных систем со своими механизмами регуляции. На уровне донора - это регуляция интенсивности фотосинтеза в хлоропластах, поступления триозофосфатов в цитоплазму и использования их на синтез сахарозы. Фотосинтез служит также источником энергии для систем активного переноса ассимилятов через мембраны (АТР для ионных насосов, субстраты для дыхания).
Ключевую роль в механизме флоэмного транспорта играет загрузка флоэмных окончаний. В основе этой загрузки в клетках флоэмы лежит работа Н+-помпы, которая активируется фитогормонами, прежде всего ауксином. Абсцизовая кислота блокирует Н+/К+ обмен. Как известно, содержание АБК возрастает в тканях листа при неблагоприятных условиях, особенно при водном дефиците.
Важнейшую роль в регуляции флоэмного транспорта играет интенсивность разгрузки флоэмы в зонах потребления ассимилятов. При ослаблении аттрагирующего действия акцепторных органов снижается продольный осмотический градиент в ситовидных трубках, а следовательно, и градиент гидростатического давления - главная движущая сила флоэмного транспорта. Высокая аттрагирующая способность акцепторных органов зависит от интенсивности их роста и активности Н+-насосов. То и другое контролируется фитогормонами. Локальное воздействие абсцизовой кислоты на флоэму облегчает отдачу ассимилятов в зонах разгрузки.
Регуляция флоэмного транспорта может осуществляться и на уровне мембранных переносчиков сахарозы. Установлено, что при повышении концентрации ассимилятов в ситовидных трубках из-за интенсивного фотосинтеза или ослабления аттрагирующей активности потребляющих органов в ситовидных трубках повышается тургорное давление, что в свою очередь ингибирует работу переносчиков сахаров. Предполагается, что при повышении тургорного давления плазмалемма в клетках флоэмных окончаний листа прижимается к клеточным стенкам, что обратимо деформирует структуру переносчика и таким образом снижает интенсивность поступления сахарозы в ситовидные трубки.
Активация транспорта ассимилятов по флоэме наблюдается при прохождении по проводящему пучку электрического импульса (потенциала действия). Флоэмный транспорт зависит от достаточного снабжения растительного организма калием, а также от температуры, так как механизм передвижения ассимилятов обусловлен работой ферментных систем (ионных насосов). Для энергетического обеспечения функциональной активности трансмембранных насосов необходим кислород.
ПЕРЕДВИЖЕНИЕ ОРГАНИЧЕСКИХ ВЕЩЕСТВ. В растениях перемещается множество органических соединений, в том числе углеводы, азотсодержащие вещества, регуляторы роста, витамины, вирусы и различные биоциды - гербициды, фунгициды и инсектициды. Большая часть органических соединений перемещается по флоэме, как показывают опыты с кольцеванием, анализы флоэмного сока и эксперименты с радиоактивными метками. Растущее растение можно рассматривать как интегральную систему потребителей питательных веществ, конкурирующих между собой в отношении направления передвижения и количества углеводов и азотсодержащих веществ, двигающихся к различным тканям в зависимости от нужд потребителей и значения разных источников снабжения. Таким образом, любое обсуждение вопроса о передвижении веществ требует количественной характеристики происходящего роста и места, где он наблюдается.
УГЛЕВОДЫ. Количество передвигающихся углеводов превышает общее количество всех других растворенных веществ. Углеводы передвигаются преимущественно вниз, но отмечается и значительное движение вверх. Их источниками служат фонды запасных углеводов и продукты текущего фотосинтеза. Количество передвигающихся веществ зависит от потребности в углеводах и близости растущей ткани к источникам снабжения. У большинства древесных растений углеводы передвигаются в виде сахарозы. У немногих семейств важными компонентами флоэмного сока являются рафиноза, стахиоза, вербаскоза. Сахароспирт сорбит обнаружен у яблони и вишни, маннит - у ясеня. Редуцирующие сахара во флоэмном соке не встречаются. Библейская "манна небесная" может происходить из разных источников, но имеющаяся в продаже манна (медвяная роса) добывается из содержащего маннит флоэмного сока ясеня манного при его подсочке.
У молодых голосеменных растений умеренной зоны до распускания почек главным потребителем углеводов являются обычно корни. После распускания почек происходит обратное передвижение, когда большее количество углеводов движется к развивающимся побегам. В это время рост камбия является второстепенным потребителем притекающих углеводов. После окончания сезонного роста побегов основными потребителями углеводов являются растущий камбий и корни, возобновляющие рост. Этот обобщенный сезонный ход событий может сильно изменяться в год обильного плодоношения, когда репродуктивные ткани монополизируют основную массу имеющихся углеводов. При одновременном росте корней и побегов соотношение остающихся в побегах и передвигающихся в корни ассимилятов зависит от относительного роста каждого из этих потребителей и их близости к источникам углеводов.
... Когда ток по флоэме блокировали кольцеванием главного ствола на уровне земли перед началом вегетационного периода, предотвращая передвижение по флоэме запасных углеводов из корней в надземные ткани, последующий рост побегов подавлялся лишь незначительно.
Имеются доказательства того, что по мере увеличения размера деревьев и удлинения пути, по которому происходит передвижение веществ от побегов к корням, растущие ткани верхушки кроны все меньше зависят от запасов углеводов, находящихся в нижней части ствола и в корнях. Таким образом, значение нижних ветвей для верхней части стебля неуклонно уменьшается по мере роста дерева.
Рост побегов. На рост побегов используются как запасные углеводы, так и продукты текущего фотосинтеза. При этом их соотношение изменяется в зависимости от вида, генетической природы, типа побегов и расположения побегов на стволе. Растущие побеги покрытосеменных растений используют продукты текущего фотосинтеза в дополнение к утилизации запасных углеводов в начале роста. У голосеменных источники вновь образующихся углеводов, используемых на рост побегов, изменяются по временам года.
Так, в начале мая, когда у побегов сосны смолистой в центральных районах Висконсина (США) раскрываются верхушечные почки, они начинают мобилизовывать углеводы, в основном продукты фотосинтеза из однолетней хвои. К 1 июня, когда побеги находятся на стадии "свечки" (междоузлия хорошо вытянуты, а хвоя вдоль междоузлий относительно недоразвита), однолетняя хвоя поставляет большую часть продуктов текущего фотосинтеза разрастающимся побегам, тогда как 2-3-летняя хвоя дает меньше. Снабжение побегов продуктами текущего фотосинтеза из старой хвои снижается в середине августа. К этому времени полностью развившаяся хвоя текущего года замещает старую хвою в качестве основной фотоассимилирующей ткани и поставщика углеводов для роста различных других тканей. Обнаружено также, что передвижение меченых ассимилятов из старой хвои к растущим побегам сосны смолистой бывает наибольшим в период быстрого удлинения новой хвои.
Старая хвоя - самый важный источник метаболитов для роста побегов сосны смолистой. Козловский и Винжет (1964) изучали роль запасных веществ ветвей, стволов и корней, а также роль резервов вместе с продуктами текущего фотосинтеза хвои для роста побегов 8-летних деревьев сосны смолистой. С этой целью исследователи удаляли листву, блокировали флоэму кольцеванием, применяли различные комбинации этих методов до начала вегетационного периода. Дефолиация и кольцевание, отдельно или вместе, уменьшали рост побегов. Рост побегов в результате этих воздействий снижался в таком порядке:
кольцевание ветвей + удаление хвои > кольцевание у основания главного ствола + удаление хвои > удаление хвои > кольцевание ветвей > кольцевание у основания ствола.
Участие в росте побегов запасных веществ (из тканей кроме старой листвы) изменялось так: ветви > главный ствол > корни.
Направление передвижения углеводов от отдельных листьев разнолистных побегов покрытосеменных растений изменяется по мере того, как побег разрастается и на нем продолжает увеличиваться количество новых листьев. Очень молодые листья получают углеводы от зрелых листьев, расположенных ниже. Развившиеся до некоторой степени листья и получают и отдают углеводы. Полностью развившиеся листья только отдают углеводы. С разрастанием листа гетерофильного побега характер передвижения углеводов изменяется от движения вверх до движения в обоих направлениях и, наконец, когда выше его оказываются несколько хорошо развитых листьев, к преимущественному движению вниз. В итоге, когда листья стареют фотосинтез и отток ассимилятов уменьшаются. Большая часть передвигающихся вверх углеводов используется на развитие сосудистых и механических тканей стебля.
Растущие плоды часто видоизменяют характер оттока углеводов из листьев разнолистных побегов. Плоды - мощные потребители углеводов, и они часто монополизируют имеющиеся углеводы до такой степени, что вместо нисходящего передвижения наблюдается двустороннее. При этом часть продуктов фотосинтеза направляется вверх, в плоды из листьев, находящихся ниже, а часть передвигается вниз от листьев, расположенных на ветви выше.
Согласно Кридеману (1969) быстрорастущие побеги апельсина получают большие количества углеводов. Но как только листья текущего цикла роста (самой близкой по времени вспышки роста) или предыдущих циклов полностью развиваются, они становятся поставщиками углеводов в основном для расположенных рядом растущих плодов. Зрелые листья, находящиеся на некотором удалении от растущего побега, поставляют углеводы преимущественно корням.
Камбиальный рост. Зона камбия - относительно диффузная меристема и слабый потребитель углеводов. Поэтому она не очень успешно конкурирует с мощными потребителями - быстро удлиняющимися побегами и корнями или разрастающимися репродуктивными структурами. Тем не менее активный камбий, покрывающий древесину главного ствола, всю систему ветвей и главные корни, потребляет за вегетационный период большие количества углеводов. Они используются на образование новых инициальных клеток камбия и на прирост ксилемы и флоэмы. Так как большая часть новых инициальных клеток отделяется путем псевдопоперечнбго деления, происходит избыточное образование и потеря многих новых клеток. Выживают инициальные клетки, имеющие лучший контакт с лучами, что свидетельствует о сильной конкуренции за углеводы и воду.
У многих лиственных видов активность камбия на участках ниже почек начинается до их раскрытия, поэтому камбиальный рост в начале вегетационного периода должен зависеть от запасов углеводов. В частности, значительные количества запасных углеводов используются на образование флоэмы, начинающееся очень рано и часто предшествующее образованию ксилемы, иногда на несколько недель (см. главу 3). Такой ранний камбиальный рост сопровождается истощением резервных углеводов в запасающих тканях. У голосеменных растений на камбиальный рост в начале вегетационного периода используются, помимо запасных углеводов, продукты текущего фотосинтеза.
Фонд углеводов для роста камбия используется неравномерно во времени и пространстве. У многих видов умеренной зоны углеводы используются на камбиальный рост в течение лишь немногих месяцев года. И даже в это время углеводы используются периодически вследствие конкуренции камбиального роста с более мощными потребителями углеводов и вследствие влияния на снабжение углеводами колебаний климатических факторов, нападения насекомых, болезней и т.д. У многих тропических видов углеводы используются на рост камбия в течение всего года, но скорость мобилизации углеводов подвержена сезонным колебаниям. Иногда тропические деревья, у которых прирост камбия происходит ежемесячно, образуют в течение года многочисленные кольца роста, что свидетельствует о неравномерной утилизации углеводов.
Использование углеводов также сильно изменяется в зависимости от высоты ствола и в разных ветвях. Например, у сеянцев и у мощных, растущих на открытом месте взрослых деревьев больше углеводов используется на камбиальный рост в нижней части ствола по сравнению с верхней. У угнетенных и очень старых деревьев утилизация углеводов при росте в основном ограничена верхней частью ствола, что подтверждается частым выпадением колец древесины в нижней части ствола.
У голосеменных деревьев мутовки ветвей верхней части кроны являются основным источником продуктов текущего фотосинтеза для камбиального роста. Например, у типичных деревьев сосны смолистой большую часть углеводов, используемых на камбиальный рост, дают ветви верхней трети кроны. Такие относительно незатененные ветви имеют высокую фотосинтезирующую способность и короткий путь передвижения углеводов в главный ствол. Относительное значение углеводов, образуемых другими ветвями, изменяется в зависимости от условий роста, возраста деревьев, конкурентных отношений и других факторов. У растущих на открытом месте деревьев сохраняющиеся на нижней части ствола ветви дают камбиальному росту этой части важный источник снабжения углеводами. В сомкнутых насаждениях в условиях жесткой конкуренции естественное опадение нижних ветвей или потеря ими мощности ограничивают источники углеводов для камбиального роста ветвей верхней части ствола. Приток углеводов для камбиального роста из отдельных ветвей уменьшается по мере их угнетения. У голосеменных, например, стареющие нижние ветви покрываются сверху все новыми и новыми мутовками ветвей, в результате они все больше затеняются и уменьшается их фотосинтетическая активность. В этих нижних, угнетенных, ветвях меньше углеводов, пригодных для оттока и использования на камбиальный рост, по сравнению с ветвями, расположенными выше на стволе.
Как отмечалось ранее, у многих взрослых деревьев нижние, угнетенные, ветви не участвуют в снабжении углеводами роста главного ствола. Такие "отрицательные" ветви могут даже не давать достаточно углеводов для образования ксилемы по всей длине самой ветви. Например, у взрослых деревьев дугласии в основании нижних ветвей часто было на 9—10 ксилемных колец меньше, чем в главном стволе у места отхождения этой ветви. Ветви сосны ладанной в нижней половине кроны, имеющие менее трех веточек, или ветви нижней четверти кроны менее чем с пятью веточками совсем не дают углеводов для роста главного ствола.
Значение разновозрастной хвои как источника углеводов для камбиального роста изменяется в зависимости от времени года. По данным Гордона и Ларсона (1968, 1969), у молодых деревьев сосны смолистой большая часть весенней древесины в старых междоузлиях стволов образовывалась во время удлинения побегов и на ранних этапах развития хвои, а новая хвоя в это время не давала заметных количеств ассимилятов на камбиальный рост. На образование весенней древесины скорее всего использовались углеводы, оттекающие из старой хвои, в основном однолетней. Новая, почти полностью развившаяся хвоя начинает давать большие количества углеводов на образование ксилемы. В это же время наблюдалось изменение направления, в котором передвигались углеводы из старой хвои: теперь транспорт происходил главным образом в корни и очень мало углеводов двигалось вверх, к новым ветвям. Усиленное снабжение углеводами после созревания новой хвои коррелировало с утолщением вторичных стенок трахеид, характерным для развития поздней древесины. Ближе к концу вегетационного периода новая хвоя снабжала углеводами новые почки и камбиальный рост верхних междоузлий ствола, а старая хвоя поставляла углеводы для роста камбия нижних междоузлий. В конце сезона, когда камбий уже не был заметным потребителем, хвоя всех возрастов снабжала углеводами корни, и в паренхимных тканях по всему дереву накапливались запасные углеводы.
Репродуктивный рост. Растущие плоды являются мощными потребителями, использующими большие количества запасных и (или) вновь образованных углеводов. Например, у бука много запасных углеводов передвигалось в развивающиеся цветки и плоды главным образом из древесины ветвей. Запасы крахмала осенью и в начале зимы явно уменьшались из-за потребностей растущих плодов. Предпочтительное использование углеводов на репродуктивный рост было показано Риго и Дависом (1959) для пяти сортов персиковых деревьев, плодоносящих в разное время. У каждого сорта содержание крахмала в плодоносящих ветвях уменьшалось перед сбором урожая плодов и увеличивалось после него. На запасы углеводов сильно влияет периодичность плодоношения. Например, содержание крахмала в листьях мандариновых деревьев, образующих немного плодов, было примерно вдвое выше, чем в листьях деревьев с обильным урожаем. Молодые и старые плоды на одном и том же дереве также конкурируют за углеводы. В случаях, когда на деревьях оставляли зрелые апельсины, в течение нескольких месяцев до сбора урожая отмечались четкая периодичность плодоношения и уменьшение размеров очередного урожая. Происходило это, вероятно, потому, что поздний сбор уменьшал количество углеводов, которыми располагает дерево для последующего урожая.
На пятую или шестую неделю после полного цветения содержание крахмала в плодоносящих плодовых веточках яблони быстро падало приблизительно до 1/3 содержания в неплодоносящих веточках. Это быстрое истощение запасов совпадает с сильным увеличением содержания гиббереллина в развивающихся семенах. Продолжительный приток гормонов из растущих семян, по-видимому, стимулирует передвижение углеводов в быстро растущие плоды.
Наряду с мобилизацией резервов, репродуктивные структуры используют также большие количества продуктов текущего фотосинтеза. В начале вегетационного периода основная масса ассимилятов поступает в вегетативные ткани, но в конце лета большая часть их передвигается в плоды. Косточковые плоды особенно интенсивно потребляют углеводы в процессе развития. Например, персики и абрикосы являются такими мощными потребителями, что они успешно конкурируют за продукты текущего фотосинтеза с расположенными рядом развивающимися листьями. Это контрастирует с ситуацией у цитрусовых, где более мощными потребителями являлись молодые, быстро растущие вегетативные ткани.
РЕГУЛЯТОРЫ РОСТА. Регуляторы роста всех классов, за исключением этилена, обнаружены как в ксилеме, так и во флоэме. Передвигаются они, по-видимому, на большие расстояния. Меченые регуляторы роста передвигались из листьев с такой же скоростью, что и другие органические соединения, а вещества, стимулирующие цветение, быстро переходят из листьев в кончики стеблей. Ауксин, гиббереллин, абсцизовая кислота и цитокинины (см. главу 15) передвигаются из корней по ксилеме. Таким образом, нет сомнений в наличии транспорта регуляторов роста на большие расстояния. Однако физиологическое значение такого дальнего транспорта неясно, так как регуляторы роста могут образовываться во всех органах растений. Поэтому представляется возможным, что синтезом in situ объясняется большинство результатов, приписываемых транспорту из корней и других отдаленных тканей. Для оценки значения транспорта на большие расстояния необходимо дальнейшее изучение источников и потребителей.
ВИРУСЫ. Вирусные инфекции возможны только при их прямом введении в определенные ткани, обычно при участии насекомого-переносчика. Все вирусы могут проникать в паренхимные клетки. Некоторые вирусы, по-видимому, адаптированы к определенным тканям, а другие (например, вирусы мозаики) могут инфицировать большинство живых клеток растения. У основной части более специализированных вирусов инфицирование зависит от флоэмной ткани, хотя некоторые, видимо, передвигаются по ксилеме.
Возбудитель вирусной инфекции перемещается по флоэме с током ассимилятов. Поскольку флоэма пронизывает все растение, а транспорт происходит в соответствии с характером изменений потребителей, вирусная инфекция может легко распространиться по всему растению. Инфекция, вызванная вирусами, передвигающимися только по флоэме, не может распространяться за окольцованную часть ствола. В противоположность этому инфекция, вызванная вирусами мозаики, может обойти кольцо через клетки паренхимы в ксилеме окольцованной части ствола. Поскольку инфекционный возбудитель вирусной болезни, передвигаясь по растению, может оставить некоторые его части неинфицированными, вполне вероятно, что возбудитель переносится пассивно. Некоторые вирусные инфекции причиняют серьезный вред флоэме, другие нет.
ЕВРАЗИЙСКИЙ НАЦИОНАЛЬНЫЙ УНИВЕРСИТЕТ ИМЕНИ Л.Н.ГУМИЛЕВА
Дата добавления: 2015-02-03; просмотров: 1101;