Участие микроорганизмов в биологическом круговороте углерода и азота
Участие микроорганизмов в биологическом круговороте углерода. Главная роль в конструктивном обмене микроорганизмов принадлежит углероду. В природе практически нет ни одного углеродсодержащего соединения, на которое не воздействовали бы микроорганизмы. По отношению к углероду все организмы делятся на автотрофные, способные синтезировать органические соединения из СО2 внешней среды, и гетеротрофные, источником углеродного питания которых является готовое органическое вещество.
Гетеротрофы делятся на паразитов и сапрофитов. Паразиты живут за счет органических веществ других организмов, а сапрофиты питаются мертвым органическим материалом. Среди паразитов различают такие, которые способны развиваться только внутри клеток хозяина (облигатные паразиты, например, хламидии, лишенные собственной системы фосфорилирования), и такие, которые способны вести сапрофитный образ жизни (факультативные паразиты). Последних значительно больше, чем первых. Сапрофиты составляют основную массу почвенных микроорганизмов. Они перерабатывают огромные массы органических веществ мертвых остатков растений и животных и содержащихся в продуктах жизнедеятельности почвенных организмов, а также в органических удобрениях.
Автотрофы, в свою очередь, делятся на фотоавтотрофов (водоросли, цианобактерии, пурпурные и зеленые бактерии) и хемоавтотрофов (нитрифицирующие, водородные, бесцветные серные и железобактерии).
Из фотоавтотрофов водоросли и цианобактерии осуществляют процесс фотосинтеза с выделением О2 подобно высшим растениям. Они имеют обе фотосистемы (ФС-1 и ФС-2). У пурпурных бактерий содержится бактериохлорофилл, активная форма которого представлена пигментом П870, а у зеленых серобактерий – П840. У пурпурных и зеленых бактерий функционирует только ФС-1, и фотосинтез происходит без выделения кислорода. Отличается эта группа и донорами протонов и электронов. Так, у зеленых и пурпурных серных бактерий таким донором является сероводород:
СО2 + 2Н2S → (СН2О)n + 2S + Н2О
У пурпурных несерных бактерий в качестве доноров протонов и электронов служит молекулярный водород или водород спиртов и некоторых других органических соединений: СО2 + 2Н2 → (СН2О)n + Н2О.
Хемоавтотрофные бактерии получают энергию для синтеза органических веществ за счет окисления различных неорганических соединений: аммиака, водорода, сероводорода, серы, закисного железа и других. Процесс синтеза органических веществ из внешней углекислоты за счет энергии окисления неорганических соединений, как указывалось выше, носит название хемосинтеза.
Большинство микроорганизмов, как и высшие растения, используют в качестве источника энергии окисление органических веществ в процессе дыхания. Для анаэробных микроорганизмов важнейшим источником энергии является сбраживание органических веществ. Различают следующие типы брожений: спиртовое; молочнокислое; маслянокислое; пропионовокислое и др.
Возбудителями спиртового брожения служат главным образом дрожжевые грибы. Пировиноградная кислота с помощью фермента пируватдекарбоксилазы расщепляется до уксусного альдегида и СО2. Образующийся ацетальдегид при участии НАД - зависимой алкогольдегидрогеназы восстанавливается до этилового спирта.
В анаэробных условиях дрожжи осуществляют спиртовое брожение. При доступе кислорода они прекращают ход спиртового брожения и начинают активно размножаться почкованием. Поэтому в анаэробных условиях получают спирт, а в аэробных - коммерческие дрожжи. Из сахаров легче всего сбраживается глюкоза, фруктоза, сахароза и мальтоза. Плохо поддаются этому процессу манноза и галактоза. Кроме дрожжей спиртовое брожение могут вызывать некоторые бактерии, в частности Zymomonas mobilis, Sarcina ventriculi и др.
Возбудителями молочнокислого брожения являются бактерии из родов Streptococcus, Lactobacillus, Pediococcus и некоторых других. Все они грамположительны, не образуют спор, не способны к передвижению и являются факультативными анаэробами. Они вызывают типичное гомоферментамивное молочнокислое брожение, конечный продукт которого - молочная кислота. При гомоферментативном молочнокислом брожении пировиноградная кислота восстанавливается с участием НАДН2 и дает молочную кислоту. Молочнокислое брожение используется для получения кисломолочных продуктов, при квашении овощей и силосовании кормов.
Другая группа молочнокислых бактерий (род Leuconostoc, некоторые виды рода Lactobacillus) в процессе гемероферменмамивного молочнокислого брожения продуцирует наряду с молочной ряд других веществ: уксусную кислоту, этиловый спирт и другие продукты. Гетероферментативное молочнокислое брожение осуществляется по окислительному пентозофосфатному пути, один из продуктов которого - ФГА превращается в ПВК, а второй - ацетилфосфат дает уксусную кислоту или этиловый спирт и СО2. Факультативно анаэробная кишечная палочка, обитающая в кишечнике животных, в анаэробных условиях сбраживает субстрат до молочной, уксусной, янтарной и муравьиной кислот, этилового спирта и молекулярного водорода.
Процесс сбраживания углеводов до масляной кислоты носит название маслянокислого брожения. Возбудителями маслянокислого брожения являются бактерии из рода Clostridium. В результате окислительного декарбоксилирования ПВК образуется ацетилКоА, который частично превращается в ацетилфосфат, а затем в уксусную кислоту. На этом этапе образуется молекула АТФ. Путь, непосредственно ведущий к синтезу масляной кислоты, начинается с конденсации двух молекул ацетил-КоА и через ряд восстановительных реакций в конечном итоге приводит к образованию масляной кислоты.
Для биологического круговорота веществ в лесу особенно важными являются две разновидности маслянокислого брожения:
· сбраживание пектиновых веществ, содержащихся в срединных пластинках клеточных стенок растений;
· анаэробное разложение целлюлозы - основного компонента оболочек растительных клеток.
Пектины состоят из довольно большого количества остатков D-галактуроновой кислоты и различных сахаров: галактозы, ксилозы и арабинозы. Разложение пектиновых веществ начинается с их гидролиза, а собственно маслянокислому брожению подвергаются только сахара - продукты гидролиза. При этом из галактозы образуются масляная кислота, углекислота и водород, а из арабинозы и ксилозы, относящихся к группе пентоз, - масляная кислота, углекислота и вода.
В анаэробных условиях, в частности в заболоченных лесных почвах, происходит медленное гидролитическое расщепление целлюлозы до глюкозы, которая затем сбраживается бактериями из рода Clostridium до масляной кислоты и ряда других продуктов.
В природе наряду с анаэробным сбраживанием существуют и аэробные процессы разложения веществ и целлюлозы, осуществляемые микроорганизмами различных систематических групп: миксобактериями, актиномицетами и грибами. В кислых лесных почвах основную роль в разложении пектиновых веществ играют грибы, в особенности мукоровые, а клетчатки - микромицеты Trichoderma, Penicillium, Aspergillus и др. Целлюлоза в почвах степных и луговых формаций разлагается в основном бактериями.
В лесных почвах в больших количествах наряду с целлюлозой находится и лигнин. Это очень медленно гидролизуемый компонент растительного спада. Находящиеся в его составе ароматические углеводороды трудно поддаются разложению. Лигнин разлагается грибами и небольшим числом бактерий, в основном из рода Pseudomonas. Из разрушающих лигнин грибов следует, прежде всего, назвать базидиальные. После частичного разрушения лигнина базидиальными грибами в работу включаются микромицеты Fusarium, Trichoderma, Rhizoctonia, Penicillium, Aspergillus и др. На самых последних стадиях разрушения лигнина участвуют и некоторые грамотрицательные бактерии. Обычно процессы гидролиза лигнина и целлюлозы в почве идут параллельно и могут вызываться одними и теми же микроорганизмами и высшими грибами.
Гидролиз лигнина сопровождается синтезом веществ, являющихся составными элементами гумуса почвы. Ароматические спирты, продукты микробиологического распада лигнина, конденсируются с аминокислотами почвы и пептидами, образуя первичные продукты гумусовых веществ. В процессе гумификации растительных остатков, в том числе содержащих лигнин, дубильные вещества, белки, смолы, воски и некоторые другие, происходит постепенное окисление гидроксильных групп до карбоксильных. Образующиеся высокомолекулярные вещества, способные осаждаться кальцием, получили название гумусовых кислот, а более дисперсная часть гумуса - фульвокислот.
Наряду с синтезом происходит и распад гумуса почвы. Продукты распада служат основой минерального питания растений. В этом процессе участвуют бактерии из родов Pseudomonas, Seliberia, Agrobacterium, а также актиномицеты и грибы. Поэтому обогащение почвы гумусом - это результат положительного баланса между образованием и разрушением гумусовых веществ.
В составе растительного опада и трупных останков животных трудно гидролизуемыми органическими веществами являются и жиры. Они разлагаются в почве аэробными бактериями из рода Pseudomonas, актиномицетами, а также анаэробными бактериями, в особенности представителями рода Clostridium.
Возбудителями пропионового брожения являются грамположительные, не образующие спор, бактерии из рода Propionibacterium. Конечный продукт брожения - пропионовая кислота. Основной начальный этап процесса данного брожения - гликолиз. Вместе с тем у данной группы бактерий обнаруживается и окислительный пентозофосфатный путь, цикл Кребса и т.д. У некоторых видов проявляется эффект Пастера. К среде эти микроорганизмы малотребовательны. Они проявляют свою активность после деятельности молочнокислых бактерий, сбраживая молочную кислоту до пропионовой и уксусной. Важно отметить, что в начале процесса ПВК не восстанавливается, как в других видах брожений, а карбоксилируется с образованием щавелевоуксусной кислоты, дальнейшее превращение которой происходит в цикле Кребса. Процесс карбоксилирования ПВК получил название гетеротрофной или темновой фиксации СО2. В небольших масштабах эта гетеротрофная фиксация СО2 наблюдается почти у всех живых организмов, в том числе у животных.
Рассмотрим схему биологического круговорота углерода. Автотрофы - высшие зеленые растения, водоросли, цианобактерии, фотосинтезирующие и хемосинтезирующие бактерии - фиксируют углерод СО2, включая его в органические соединения. Животные, поедая растения, концентрируют углерод в органическом веществе своего тела. Дыхание аэробных организмов и многие виды брожений сопровождаются выделением углекислоты. Опад растений и трупные останки животных и микроорганизмов разлагаются и пополняют атмосферу СО2. Образующаяся углекислота опять используется в процессах фото- и хемосинтеза. Цикл биологического круговорота углерода повторяется снова. В этом круговороте прослеживается тесная связь жизнедеятельности растений, животных и микроорганизмов.
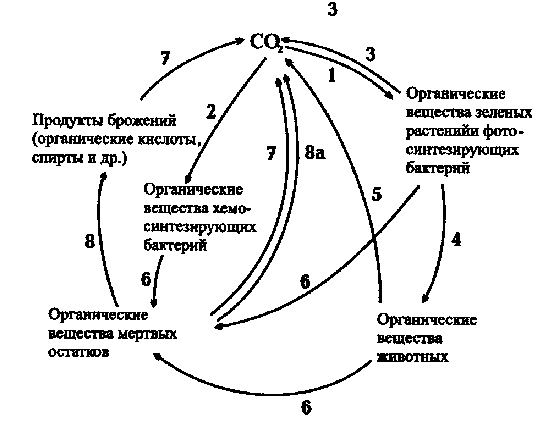
Схема биологического круговорота углерода: 1 - фотосинтез; 2 - хемосинтез; 3 - дыхание растений; 4 - питание животных; 5 - дыхание животных; 6 - отмирание организмов и их прижизненные выделения; 7 - дыхание аэробных микроорганизмов; 8 - брожение; 8а - выделение СО2 при брожениях
Участие микроорганизмов в биологическом круговороте азота. Наряду с участием в биологическом круговороте углерода микроорганизмы выполняют очень важную роль в преобразовании азотистых веществ. Это значение микробов трудно переоценить, ибо недостаток в почве важнейшего мя растений элемента азота обычное явление. Азотное питание в сельском и лесном хозяйстве представляет собой проблему номер один.
Азот, как и углерод, является основным компонентом наиболее существенных для жизни растений органических веществ. После отмирания растений, микроорганизмов, животных органическое вещество их опада и трупных остатков разлагается почвенными микробами до самых простых минеральных форм азота, так нужных растениям. Это происходит с помощью большой группы микроорганизмов - аммонификаторов, а процесс по конечному продукту (аммонию) получил название аммонификации.
В аммонификацию включаются белки, мочевина, хитин, нуклеиновые кислоты - вещества, имеющие большой запас азота. Все они активно атакуются аммонифицирующими микроорганизмами с образованием аммиака. Разлагая то или иное азотсодержащее органическое вещество, возбудители рассматриваемого процесса за счет этого питаются, размножаются, черпают необходимое количество энергии для своей жизнедеятельности.
Обычно микробы при этом используют незначительную долю высвобождающегося азота на поддержание собственного метаболизма. Большая часть подвижного и хорошо доступного корням азота остается в почве, что благоприятно сказывается на азотном питании растений и на их урожайности. Одновременно при разложении органических веществ почвенными микроорганизмами высвобождаются ионы ортофосфорной кислоты, а также сера в форме H2S; последний окисляется серными бактериями до сульфатов.
Возбудителями аммонификации белков являются, с одной стороны, аэробы - спорообразующие палочки Bacillus mycoides, В. megaterium, В. subtilis (сенная палочка), многие актиномицеты и грибы, а с другой анаэробные палочки из рода клостридиум (С. putrificum, С. sporogenes, С. botulinum, С. tetani). Два последних вида очень вредны для человека: первый вызывает пищевое отравление вплоть до летальных последствий, а второй - возбудитель столбняка.
Все названные аммонификаторы, или гнилостные микроорганизмы, вырабатывают и выделяют большое количество протеолитических ферментов - протеаз, осуществляющих гидролиз белков. Процесс распадается на два этапа.
На первом этапе с помощью протеаз сложные белки разлагаются до простых белков, а те, в свою очередь, до аминокислот. Образующиеся аминокислоты частично усваиваются микробами, из них они строят белки своего тела. В этом случае схема реакций дезаминирования аминокислот выглядит так:
R-CHNH2-COOH + О2 → R-COOH+CО2 + NН3 или
R - CHNH2 - СООН + 1/2 О2 → R - СО - СООН + NН3.
Большая часть аминокислот на втором этапе подвергается дезаминированию, в процессе которого от них отщепляется аминогруппа NH2 и образуются свободный аммиак и органические кислоты. Различают дезаминирование, идущее с участием и без участия кислорода. Процесс осуществляется с помощью НАДзависимой дегидрогеназы:
R - CHNH2 - СООН + НАД + Н2О → R - СО - СООН + НАД-Н2 + NН3.
Конечные продукты аммонификации белков частично используются бактериями: аммиак - в конструктивном обмене в качестве источника азота, а органические кислоты - в энергетическом метаболизме.
Благоприятными внешними условиями для аммонификации белков являются температура в пределах 20 - 30° С, достаточное количество воды, нейтральная или щелочная среда (рН 7 и выше).
Мочевина является продуктом жизнедеятельности многих грибов и животных. В чистом виде она труднодоступна корням растений. Аммонификаторы разлагают ее до аммиака, предоставляя растению удобную форму азотного питания. К уробактериям, осуществляющим аммонификацию мочевины, относятся Micrococcus urea, Sporosarcina urea, Bacillus pasteurii и др. Они вырабатывают специальный фермент уреазу и, выделяя его в среду, осуществляют рассматриваемый процесс. Аммонификация мочевины состоит из двух этапов: сначала происходит гидролитическое дезаминирование мочевины с образованием углекислого аммония:
CO(NH2)2 + Н2О → (NН4)2СО3;
затем углекислый аммоний как вещество очень нестойкое распадается на аммиак, углекислоту и воду:
(NН4)2СО3 → 2 NН3 + СО2 + Н2О.
Образующийся аммиак частично участвует в конструктивном обмене уробактерий, а в остальном пополняет запасы доступного для растений азота в почве.
Очень сложным и трудно разлагаемым в почве азотсодержащим веществом является хитин. Он входит в состав клеточных стенок грибов, наружного скелета ракообразных и насекомых. В химическом отношении является аналогом целлюлозы, лишь одна гидроксильная группа глюкозы заменяется группой СН3-СО-NН-. Элементарное звено молекулы этого азотсодержащего полисахарида представлено ацетилглюкозамином.
Возбудителями процесса аммонификации хитина являются бактерии из родов Bacillus, Pseudomonas, Achromobacter, а также плесневые грибы, главным образом из рода Aspergillus, и актиномицеты:
· на первом этапе с помощью эктоферментов микроорганизмы расщепляют хитин на его элементарные звенья - молекулы ацетилглюкозамина;
· ацетилглюкозамин уже на втором этапе распадается на глюкозу, уксусную кислоту и аммиак. Здесь, так же как и в предыдущих случаях, образующийся аммиак используется аммонификаторами и пополняет запасы азота в почве.
Наиболее активным возбудителем аммонификации нуклеиновых кислот является Bacillus megaterium:
· с участием синтезируемых бактериями внеклеточных ферментов (рибонуклеаза и дезоксирибонуклеаза) производится гидролиз ДНК и РНК до нуклеотидов;
· затем нуклеотиды распадаются до азотистых оснований, сахара и фосфорной кислоты;
· азотистые основания усваиваются бактериями и внутри их клеток разлагаются до аммония, углекислоты, муравьиной, уксусной и молочной кислот. Образующийся аммоний используется в качестве источника азотного питания бактерии.
Процессам аммонификации подвергаются также гуминовые кислоты, мочевая кислота и некоторые другие вещества почвы. Образующийся в процессе аммонификации перечисленных азотсодержащих соединений аммиак соединяется в почве с анионами и образует аммонийные соли - углекислые, сернокислые и др. После отмирания аммонифицирующих бактерий азот их органических соединений под действием живых аммонификаторов также пополняет почву аммонием.
Другим важным в биологическом круговороте азота процессом является нитрификация- окисление образующегося при аммонификации аммиака до азотистой и азотной кислоты. Осуществляют этот процесс бактерии из группы хемоавтотрофов.
Протекает процесс нитрификации в 2 этапа:
1. Аммиак окисляется до азотистой кислоты:
2NН3+3О2 → 2HNО2 + 2Н2О + энергия.
Этот этап осуществляют мелкие палочковидные бактерии со жгутиками из рода Nitrosomonas, а также представители родов Nitrosococcus и Nitrosospira.
2. Азотистая кислота окисляется до азотной кислоты:
2HNО2 + О2 → 2НNО3 + энергия.
Окисление происходит благодаря деятельности бактерий из родов Nitrobacter и Nitrococcus.
Высвобождающаяся при окислении аммиака и нитратов энергия идет на хемосинтез - образование из СО2 органического вещества, а также на некоторые другие процессы жизнедеятельности.
Азотная кислота, взаимодействуя с различными солями и щелочами почвы, образует нитраты - второй прекрасный источник азотного питания растений. Кроме того, азотная кислота переводит нерастворимые фосфорные соединения почвы в легкодоступные для растений формы.
Нитрификации мы обязаны образованием залежей селитры. Такие залежи возникли в Средней Азии, на Памире, в Индии, Чили, в ряде районов Африки и других местах. Наиболее богатые месторождения селитры приурочены к жаркому сухому климату и высоким горам. Особенно благоприятные условия нитрификаторы находят для себя на склонах гор.
Дождевые воды или воды вечных снегов и льдов смывают нитраты к подножию, вода здесь быстро испаряется, а нитраты накапливаются в виде больших залежей.
Нитрификация может происходить только при хорошей аэрации и нейтральной или слабощелочной реакции почвы. Достаточно активно процесс нитрификации протекает в черноземных и некоторых других почвах. В большинстве лесных почв нитрификация отсутствует или проявляется весьма слабо.
Противоположным нитрификации процессом является денитрификация,при которой происходит восстановление нитратов до молекулярного азота. Промежуточными веществами являются нитриты (NО2-), окись азота (NO) и закись азота (N2О).
Возбудителями денитрификации являются факультативно анаэробные бактерии из родов Pseudomonas, Micrococcus, в частности М. denitrificanas, и др. Процесс осуществляется ими с помощью соответствующих ферментов - редуктаз.
Общее уравнение процесса денитрификации выглядит так:
С6Н12О6 + 4NО3- → 6СО2 + 6Н2О + 2N2 + энергия.
Денитрификация происходит в условиях плотных и затопленных почв с плохой аэрацией и является весьма вредным процессом, так как образующийся молекулярный азот уходит в атмосферу и не используется растениями. В практике лесного и сельского хозяйства нельзя допускать уплотнения пахотных или лесокультурных почв, а затапливаемые почвы следует осушать. В природных лесах необходимо проводить мероприятия по регулированию наплыва городского населения с тем, чтобы не уплотнять почву до предельных величин. При хорошей аэрации денитрификация прекращается, так как возбудители этого процесса переходят от анаэробного нитратного дыхания к обычному аэробному дыханию, используя в качестве акцептора электронов молекулярный кислород.
Особо важное место в биологическом круговороте веществ в природе занимает фиксация микроорганизмами атмосферного газообразного азота. Главное значение азотфиксации сводится к вовлечению инертного молекулярного азота атмосферы в биологический круговорот, к обогащению им почвы как среды обитания растений.
Микроорганизмы, способные фиксировать молекулярный азот, называются азотфиксирующими или азотфиксаторами. Среди них различают две большие группы: свободно живущие в почве азотфиксаторы и симбионты, осуществляющие этот процесс в симбиозе с растениями. К первой группе относятся Clostridium pasteurianum и некоторые другие представители рода Clostridium, Azotobacter, Beijerinckia, многие фототрофные бактерии и цианобактерии.
Clostridiut pasteurianum - анаэробная палочка; в момент спорообразования принимающая вид веретена. Это возбудитель маслянокислого брожения. Энергия, высвобождающаяся в процессе брожения, используется этим микроорганизмом на фиксацию молекулярного азота и перевод его в состав органических веществ своего тела; на 1 г сбраживаемого сахара связывается 23 мг азота. Clostridium - строгий анаэроб, кислород является для него ядом. Вместе с тем эта бактерия широко распространена во многих почвах, в том числе хорошо дренированных, с наличием больших количеств кислорода. Объяснение заключается в том, что данный вид поселяется внутри почвенных частичек, тогда как аэробные микроорганизмы - на их поверхности. Последние, поглощая кислород, как бы фильтруют проходящий внутрь почвенного комочка воздух, лишая его кислорода.
Аэробный азотфиксатор Azotobacter chroococcum в молодых культурах представлен подвижными палочками, затем жгутики утрачиваются, клетки становятся шаровидными, обычно соединенными по 2 - 3 и окруженными слизистой капсулой. Энергию для азотфиксации азотобактер черпает за счет обычного дыхательного распада окисляемых веществ, что определяет и эффективность процесса: на 1 г расходуемого сахара фиксируется до 20 мг азота. Чистые культуры азотобактера широко используются в качестве бактериального удобрения под названием азотобактерин.
Клубеньковые бактерии способны фиксировать азот атмосферы только в симбиозе с растениями. Бактерии, живущие в клубеньках на корнях бобовых, относятся к роду Rhizobiuт. Название вида клубеньковых бактерий происходит от названия вида растений, с которыми бактерии сожительствуют. Например, если на клевере, то это будет R. trifolii, на белой акации R. robinii и т.д.
Клубеньковые бактерии живут в почве, но к фиксации азота в таком состоянии не способны. Этот процесс они могут осуществлять лишь находясь в тесном симбиотическом сожительстве, так как для азотфиксации нужна энергия, поставляемая растением-хозяином. Известно более 1300 видов бобовых растений, в корнях которых функционирует процесс азотфиксации с помощью клубеньковых бактерий. Последние представляют собой грамположительные палочки длиной всего лишь около 3 мкм. Бактерии заражают растение сразу же после прорастания семян бобовых растений, когда корень проростка проникает в почву.
Корневые волоски бобовых растений при соприкосновении с клубеньковыми бактериями выделяют особые вещества - лектины белковой природы, а бактерии - полисахариды. Под влиянием этих веществ между корневым волоском и бактериями происходит тесное взаимодействие. Корневой волосок изгибается, принимает форму ручки зонтика и начинает выделять полигалактуроназу. Этот фермент разрушает стенку своей клетки и открывает путь бактериям внутрь. Это начало пути инфекции. Бактерии в корневом волоске активно делятся, образуют инфекционную нить. Попадая в клетки коры корня, бактерии размножаются и изменяют форму: из палочек превращаются в клетки колбовидной формы - бактероиды. В дальнейшем под воздействием гормонов происходит активное деление корня с образованием клубенька.
Клубенек постепенно из белого превращается в розовый благодаря образованию леггемоглобина - белка очень сходного с гемоглобином крови животных. В этот момент устанавливается тесная функциональная связь бактериального комплекса с корнем растения, а весь комплекс представляет теперь собой азотфиксирующую единицу.
Клубеньковые бактерии не только обеспечивают азотом себя, но и до 75 % зафиксированного ими азота отдают растению-хозяину. За один вегетационный период бобовые растения фиксируют от 100 до 400 кг азота на 1 га поля. В связи с этим урожаи зерновых, посеянных вслед за бобовыми растениями, удваиваются.
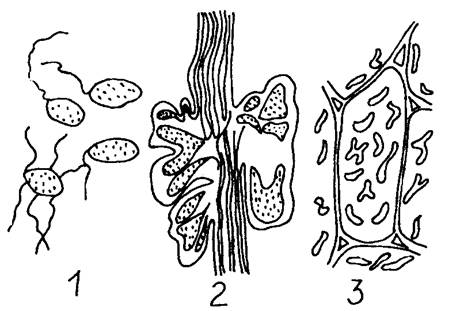
Клубеньковые бактерии: 1 – бактерии в почве; 2 – продольный разрез через корень и клубенек бобового растения; 3 – бактероиды в клетках корня
В симбиоз с азотфиксирующими микроорганизмами вступают не только бобовые, но и представители некоторых других семейств. Из древесных небобовых растений на корнях образуют клубеньки лох, облепиха, ольха. У них клубеньки формируют актиномицеты, относящиеся к роду Fraпkia. Они также способны фиксировать азот атмосферы. У некоторых древесных растений тропиков клубеньки образуются на листьях.
Механизм азотфиксации очень сложен. Наиболее трудным для азотфиксаторов является процесс разрыва первой из трех связей в молекуле азота N = = N. На это идет большое количество энергии, а именно 941 кДж/моль.
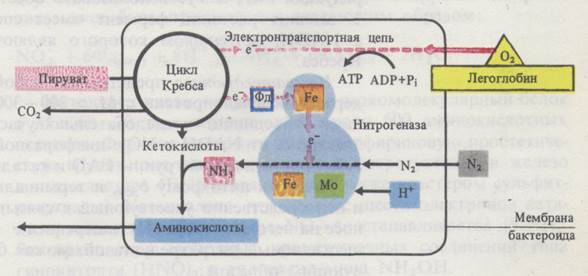
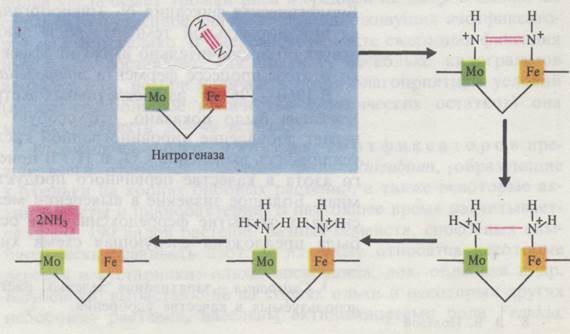
Связывание молекулярного азота осуществляется с помощью нитрогеназы - ферментного комплекса, способного активировать N2, а затем восстанавливать его. В состав нитрогеназы входят два белка: 1) азоферредоксин, содержащий железо; 2) молибдоферредоксин, содержащий два атома молибдена и 30 атомов железа. Биосинтез ферментного комплекса нитрогеназы осуществляется с участием особой плазмиды, внехромосомной ДНК. Прежде чем начинается процесс фиксации, азот активируется молибденом нитрогеназы. Железо нитрогеназы используется в качестве переносчика электронов.
Источником водорода и электронов для восстановления азота у большинства азотфиксаторов служит пировиноградная кислота. Электроны попарно передаются сначала на азоферредоксин, затем - на молибден молибдоферредоксина и, наконец, на азот. В качестве промежуточных продуктов аэотфиксации образуются диимид (NH = NН) и гидразин (H2N – NН2). Вместе с тем они могут быть и тесно связанными с ферментным комплексом - нитрогеназой. Реакции идут в следующей последовательности:
Азот → диимид → гидразин → аммиак
На каждую из этих реакций расходуется 4 молекулы АТФ, а всего на фиксацию молекулы N2 -12 АТФ. Источником АТФ является окислительное или фотосинтетическое фосфорилирование.
Данный механизм азотфиксации характерен как для свободно живущих азотфиксирующих бактерий, так и для симбионтов. Образующийся NН3 служит затем основным компонентом для синтеза аминокислот, образованию которых способствует наличие в теле азотфиксаторов большого набора органических кислот.
У клубеньковых бактерий леггемоглобин принимает участие в переносе кислорода для дыхания бактероида. Энергия для восстановления азота до аммиака поступает за счет макроэргов АТФ, образующегося в митохондриях клеток корня хозяина.
Биологический круговорот азота в природе, в котором принимают участие растения, микроорганизмы и животные, складывается из следующих процессов:
1. растения усваивают соли аммония и нитраты, включая азот этих солей в свои органические вещества; они же усваивают часть азота, связанного клубеньковыми бактериями;
2. животные, поедая растения, переводят азот в состав своих органических веществ;
3. после отмирания растений, животных и микроорганизмов трупные останки их разлагаются аммонификаторами с образованием минерального азота - аммония; часть NН3 окисляется до нитратов в процессе нитрификации, вследствие чего происходит накопление в почве нитратов;
4. определенная часть нитратов, обычно в неблагоприятных условиях аэрации, в процессе денитрификации восстанавливается до молекулярного азота N2, который улетучивается в атмосферу; почва обедняется азотом;
5. свободноживущие и симбиотические азотфиксирующие микроорганизмы связывают молекулярный азот, вовлекая его в биологический круговорот.
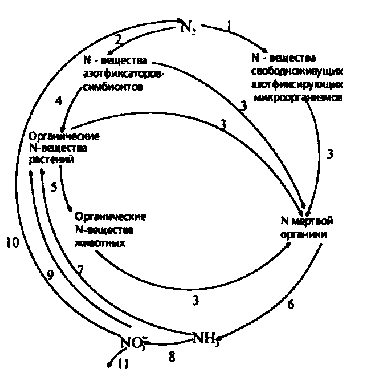
Схема биологического круговорота азота: 1 - фиксация азота свободноживущими микроорганизмами; 2 симбиотическая фиксация азота; 3 - прижизненные выделения азотсодержащих соединений, опад растений, трупные останки животных и микроорганизмов; 4 - усвоение растениями азота, связанного азотфиксаторами-симбионтами; 5 - питание животных; 6 - аммонификация; 7 - усвоение растениями аммонийных солей; 8 - нитрификация; 9 - усвоение растениями нитратов; 10 - денитрификация; 11- вымывание NО3- из воды
Дата добавления: 2015-02-03; просмотров: 11173;