НИСХОДЯЩИЕ СИСТЕМЫ ДВИГАТЕЛЬНОГО КОНТРОЛЯ
Физиология нисходящих путей от коры больших полушария.
В эволюции головного мозга заметно увеличивается площадь коры больших полушарий. В результате у высших млекопитающих, включая приматов, развивается плащ, который полностью покрывает большие полушария. Поля коры, расположенные рострально относительно центральной (у приматов) или крестовидной (у хищных) борозд, участвуют в контроле двигательной функции. Здесь локализованы центры движения глаз, моторики речи, а также центры, обеспечивающие движения конечностей и туловища.
В 1870 г. немецкие исследователи Фрич и Гитциг методом электрической стимуляции мозга собак впервые определили границы моторной коры. По современным данным, первичная моторная кора, управляющая движениями туловища и конечностей, соответствует полям 4 и 6 (по Бродману) (см. приложение 1). Ростральнее находятся корковые поля, управляющие движениями глаз (поля 8, 10) и речи (поля 44, 46). Анатомическим признаком моторных областей коры является агранулярность (невыраженность IV слоя коры). Кроме первичной моторной коры, в двигательном контроле у высших млекопитающих участвуют также премоторная, дополнительная моторная области коры, а также поле 5 (ростральная теменная кора).
Нисходящие пути двигательного контроля млекопитающих в соответствии с организацией их спинальных механизмов принято делить на латеральные (кортико- и руброспинальные) и медиальные (вестибуло- и ретикулоспинальные) проводящие системы. Латеральные проводящие системы в филогенезе млекопитающих возникли позже медиальных и обслуживают преимущественно движения конечностей, в том числе связанные с локомоцией. В отличие от этого медиальные системы преимущественно имеют отношение к рефлексам позы и установки головы и тела в пространстве.
Кортикостналъная система двигательного контроля. Пирамидный (кортикоспинальный) тракт у млекопитающих — филогенетически самый молодой двигательный тракт. Он образован аксонами крупных нейронов, тела которых находятся в пятом слое 4-го и 6-го полей коры, У обезьян большая часть волокон берет начало от 4-го (31%) и 6-го (29%) полей коры, но 40% начинаются от теменной коры. Точные данные для человека отсутствуют, но известно, что не менее 60% волокон пирамидного тракта берут начало от поля 4. У человека пирамидный тракт одной стороны содержит более 1 млн волокон.
На спинальном уровне у млекопитающих описано два кортикоспинальных тракта: мощный латеральный пирамидный тракт, волокна которого полностью перекрещиваются на уровне первого шейного сегмента, и неперекрещивающийся вентральный пирамидный тракт. Морфологические исследования показали, что у кошек н обезьян вентральный тракт заканчивается преимущественно в грудных сегментах спинного мозга. Этот тракт изучен недостаточно. Волокна латерального пирамидного тракта прослеживаются до поясничных сегментов спинного мозга. Согласно данным электрофизиологии латеральный пирамидный тракт активирует нейроны серого вещества основания заднего рога спинного мозга, которые, в свою очередь, активируют мотонейроны вентрального рога. Только у животных с подвижными фалангами пальцев (приматы, крыса, енот и некоторые другие) волокна пирамидного
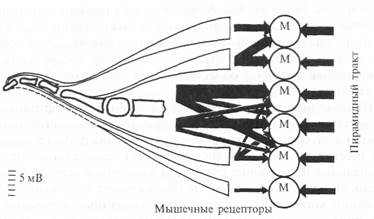
Рис. 5.20. Сравнение (показано толщиной стрелок) возбудительных входов от моторной коры и афферентов от мышечных веретен на мотонейроны спинного мозга, иннервирующие мышцы руки обезьяны.
тракта активируют эти мотонейроны непосредственно.
Экспериментально установлено, что мотонейроны, имеющие моносинаптические возбудительные связи с моторной корой, в то же время подвержены активации со стороны первичных окончаний мышечных веретен (волокна группы 1а) (рис. 5.20). По мнению исследователей, это может указывать на то, что эти мотонейроны наряду с прямым входом от моторной коры могут вовлекаться также в активность афферентов от мышечных веретен. При этом гамма-мотонейроны могут активироваться не только из моторной коры больших полушарий, но и из других головного мозга — мозжечка, красного ядра, стриатума и пр.
Электрическая стимуляция волокон пирамидного тракта в опытах на животных (кошки, обезьяны) вызывает преимущественно флексию соответствующей конечности. Это находится в полном соответствии с результатами изучения отдельных мотонейронов спинного мозга: флексорные мотонейроны на стимуляцию пирамидного тракта реагируют деполяризацией, а экстензорные — гиперполяризацией или ответом смешанного типа (де- и гиперполяризацией). При этом влияния, распространяющиеся по пирамидному тракту, носят фазический, а не тонический характер, но порог активации гамма-мотонейронов, как правило, ниже, чем альфа-мотонейронов. Корковые проекции на альфа- и гамма-мотонейроны соответствующей мышцы совпадают.
В первичной моторной коре полностью представлена соматическая мускулатура. Эффекты локальной электрической стимуляции проявляются преимущественно в активации соответствующих флексорных мышц. Более тонкие микроэлектродные исследования показали, что активацию пула мотонейронов, иннервирующих определенную мышцу, можно получить из довольно ограниченной области коры — корковой колонки. Такая колонка, например в моторной коре обезьяны, имеет площадь от 3 до 7 мм2. Как правило, в ответ на электрическую стимуляцию коры ВПСП на мембране мотонейрона нарастал ступенчато, что могло указывать на полисинаптическое проведение возбуждения к нему. Это полностью соответствует и нейроанатомическим данным, согласно которым только около 7% (у приматов) волокон кортикоспинального тракта образуют моносинаптические контакты на мотонейронах, остальные проводят возбуждение к ним через интернейроны (полисинаптически).
Современные знания о нейронной организации моторных полей коры больших полушарий в значительной степени основаны на представлении о колончатом (модульном) строении коры. На основании исследований тонкой анатомии коры было выдвинуто предположение о наличии в ней вертикальных объединений нейронов. Это было подтверждено также в физиологических экспериментах: было показано, что отдельные нейроны в такой колонке (диаметром 0,4-1 мм) имеют рецептивные поля в одном районе, например на кожной поверхности руки. Эти результаты были в дальнейшем подтверждены методом микростимуляции. Установлено, что, активируя у кошки и обезьяны небольшой участок коры электрическим током (стимуляцию производили через металлический внутрикорковый микроэлектрод), можно определить низкопороговую область активации отдельной мышцы. С помощью этого было показано, что эфферентные зоны, активирующие различные мышцы данного сустава конечности, сгруппированы в данной корковой колонке и могут частично перекрываться. Есть основания считать, что такая колонка функционирует как единица коркового моторного выхода, по крайней мере для мускулатуры дистальных (фаланги пальцев) частей конечности. Для аксиальных мышц (мышцы туловища) подобные колонки не имеют столь же четко очерченных границ, и каждая мышца может быть многократно представлена в различных корковых колонках.
Эффекты разрушений моторной коры. Локальное повреждение первичной моторной коры у обезьян вызывает вялый паралич соответствующих частей тела, однако через 1-2 нед движения в мышцах восстанавливаются, за исключением мышц фалангов пальцев, которые остаются спастичными. Эти мышцы гиперактивны, что проявляется в активации рефлексов на растяжение; наиболее выражены изменения в мышцах запястья и пальцев.
После экстирпации у обезьяны коркового представительства руки наступает ее полный паралич. Спустя месяц восстанавливается реакция схватывания, но она затянута, а мускулатура пальцев спастична. Удаление у обезьяны всей моторной коры вызывает паралич и спастичность с гипертонией флексоров рук и ног. У человека после подобных повреждений проявляется положительный рефлекс Бабинского (рефлекторное сгибание стопы при проведении черты поперек подошвы). Повреждение первичной моторной коры ухудшает независимое движение суставов. Например, пальцы у больного двигаются все вместе (рука-лопата). Обезьяна при пирамидных повреждениях не способна достать указательным пальцем изюм из узкой щели потому, что не может двигать отдельными фалангами пальцев.
Четвероногие (кошки, собаки) сразу после удаления моторной коры не могут стоять и ходить, у них исчезают реакция постановки лапы на опору* и способность переступать по движущейся относительно животного поверхности, например по ленте тредбана. Примерно на протяжении месяца после операции ходьба восстанавливается, однако специальные тесты позволяют выявить стойкие нарушения движений. Например, собака с поврежденной пирамидной системой испытывает трудности при ходьбе по гладкой (зеркальной) поверхности (лапы скользят, перекрещиваются, и животное часто падает), а крысы и кошки с большим трудом достают приманку из узкой трубки или щели. Все это указывает на нарушение тонких движений дистальных частей передних конечностей.
Сенсорные входы в моторную кору. Главный таламический вход в моторную кору позвоночных (хищных, приматов) идет от ядер
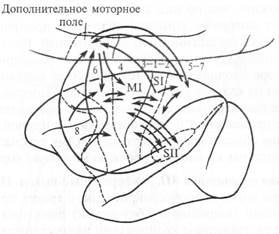
Рис. 5.21. Основные связи между корковыми областями, участвующими в моторном контроле.
Цифры обозначают поля по Бродману, стрелки показывают направление главных связей; MI - первичная моторная кора, SI — первичная сенсорная кора; SII — вторичная сенсорная кора.
вентробазального комплекса таламуса. У ненаркотизированных кошек и обезьян около 2/3 нейронов моторной коры активируются прикосновением к шерсти, пальпацией мышц и пассивными движениями в суставах; небольшая часть нейронов реагирует также на зрительные и слуховые стимулы. Нейроны в одной вертикальной проходке микроэлектрода реагируют на стимуляцию одной сенсорной модальности: например, получают вход от ограниченного участка кожи, но рецептивные поля отдельных нейронов сильно перекрываются.
Симметричные области моторной коры двух полушарий связаны между собой через мозолистое тело. Взаимными связями через мозолистое тело связаны между собой проксимальные части конечностей и аксиальная мускулатура туловища и лица. Корковые представительства дистальной мускулатуры конечностей не связаны между собой через мозолистое тело. Предполагают, что эти связи обеспечивают необходимую интеграцию двигательного контроля. Вместе с тем перерезка мозолистого тела не вызывает грубых нарушений координации движений.
Как видно из приведенной схемы на рис. 5.21, все эти связи — реципрокные (взаимные) и организованы соматотопически. Можно предположить, что по ним сенсорная (постцентральная) кора обеспечивает моторные команды кожной и проприоцептивной соматотопически организованной информацией. Вместе с тем известно, что после удаления постцентральной (сенсорной) коры в моторной коре продолжают регистрироваться вызванные потенциалы в ответ на кожные и проприоцептивные стимулы. Кроме связей MI с постцентральной сенсорной корой, она имеет реципрокную связь с премоторной и дополнительной моторной корой. Эти две последние области коры в настоящее время рассматривают как источник моторных команд в первичную моторную кору MI.
Нейронная организация MI. Эфферентный выход. Изучение активности отдельных нейронов пирамидного тракта при выполнении животными (кошками и обезьянами) движений позволило составить представление о кодировании параметров движения частотой разрядов. Было установлено, что активация пирамидного нейрона опережает активацию соответствующей мышцы (по активации ЭМГ) на 100 мс и более. Была также выявлена зависимость частоты разрядов данного пирамидного нейрона от силы (F), развиваемой мышцей, или от ее изменения (dF/dt).
Сенсомоторная организация отдельных колонок моторной коры.
Как уже указывалось выше, методом микростимуляций в моторной коре млекопитающих выявляются эфферентные колонки нейронов, которые активируют мотонейроны, принадлежащие данной мышце. Было изучено соотношение афферентных проекций и эфферентного выхода для отдельных колонок моторной коры. В качестве конкретного примера приведем результаты исследования нейронного представительства области большого пальца руки обезьяны (рис. 5.22). В области коркового представительства большого пальца производили микроэлектродную регистрацию активности отдельных нейронов и определяли их рецептивные поля. Этот же микроэлектрод использовали для электрической микростимуляции (током силой 5-10 мкА) при изучении эфферентного выхода данного нейрона (или группы нейронов, так как при микростимуляции через полисинаптические связи была задействована группа нейронов). На рис. 5.22 видно, что эфферентные нейроны данной колонки получают тактильный вход главным образом от рецептивных полей той же части пальца, которая расположена на пути его движения, вызванного внутрикорковой микростимуляцией. Специальные исследования показали, что наряду с тактильным
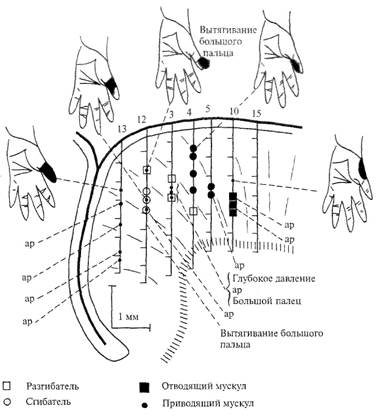
Puc. 5.22. Отношение вход-выход нейронов моторного представительства большого пальца руки обезьяны. Реконструкция путей погружения электродов и локализации нейронов одного эксперимента. Сплошные линии, обозначенные номерами, — треки электродов. Периферический двигательный эффект обозначен разными символами. Пункты, где микростимуляция вызывала двигательный эффект, обозначены короткими горизонтальными линиями на электродных треках.
входом корковые нейроны соответствующей колонки получают также информацию от суставных и мышечных рецепторов.
Было установлено, что наиболее интенсивный афферентный вход получают нейроны верхних слоев коры, а нейроны V—VI слоев, где отмечается самый низкий порог двигательных реакций в ответ на микростимуляцию, практически не получают афферентного входа. На основании этих наблюдений пришли к заключению, что в физиологических условиях афферентное возбуждение поступает на нейроны верхних слоев, соответствующим образом обрабатывается, передается на нейроны нижних слоев и там происходит формирование двигательной команды.
Участие премоторных и теменных полей коры в двигательном контроле. Корковый уровень двигательного контроля представлен также премоторными полями коры и дополнительным моторным полем, а также полем 5 теменной коры, о функции которого пока известно очень мало.
Дополнительная моторная кора (ДМК). У приматов ДМК расположена на медиальной поверхности полушария (см. рис. 5.21). Подобно первичной моторной коре ДМК организована соматотопически. Движения, вызванные электрической стимуляцией ДМК, по сравнению со стимуляцией моторной коры выглядят более сложными и растянутыми во времени. Такие движения часто похожи на целенаправленные действия, в ряде случаев они развиваются билатерально на обеих конечностях и часто длятся больше времени электрической стимуляции ДМК.
При электрической стимуляции ДМК мозга человека во время нейрохирургических операций часто наблюдали вокализацию, которая могла сопровождаться гримасами лица, координированными движениями конечностей и торможением произвольных движений.
Экстирпация ДМК у человека вызывает временную потерю речи (афазию), которая обычно проходит через несколько недель. Отмечают также замедление выполнения ритмических движений. Удаление ДМК у низших обезьян вызывает признаки нарушения целенаправленных движений (апраксию), нарушается также координация движений обеих рук. Вследствие этого у обезьян резко нарушается выполнение бимануальных задач, в которых необходима координация обеих рук. Нейронная активность ДМК коррелирует с характером движения, но установить связь с отдельными параметрами самого движения не удается. Предполагают, что нейронные сети ДМК принимают непосредственное участие в обработке сенсорной информации и в подготовке к движению (препрограммирование). В пользу этого говорит также усиление локального кровотока в ДМК у человека при намерении выполнить движение.
Участие в программировании движений премоторных полей коры. Поражение премоторных зон у человека не сопровождается параличом

Puc. 5.23. Двигательные персеверации у больного с повреждениями в премоторной области (по А. Р. Лурия, 1973).
контралатеральных конечностей. Вместе с тем их основным симптомом является отчетливое нарушение двигательных навыков. Клинически это проявляется в том, что у больного изменяется почерк, машинистка теряет быстроту и плавность своей работы, квалифицированный рабочий не способен автоматизированно выполнять серию операций, входящих в привычный двигательный акт. В клинике такие симптомы описывают под названием «инертность двигательных стереотипов». Характерными являются невозможность больного переключаться, например, с сильных ударов на слабые при отстукивании ритма ладонью, а также своеобразное «заклинивание» движения, двигательная персеверация (рис. 5.23). Наиболее отчетливо нарушения проявляются в контралатеральной очагу руке, но при поражении левого (доминантного) полушария они часто проявляются в работе обеих рук. Клиницисты отмечают, что у таких больных сохранены как намерения выполнить движение, так и общий план его выполнения, однако в своей исполнительной части движение высвобождается из-под влияния двигательной программы.
В тех случаях, когда поражение располагается в нижних отделах премоторной зоны левого (доминантного по речи у правшей) полушария, происходит нарушение речи. Нарушения проявляются при переходе больного от одной артикуляции к другой (при произнесении любого слова или словосочетания). Нарушение кинетической организации артикуляции является основой той формы моторной афазии, которую описал П. Брока (1861). Следует отметить, что нарушения у больных с поражениями нижних отделов левой премоторной зоны проявляются не только в устной речи, но и в письме. Нарушения письменной речи считают следствием нарушения плавности перехода от одного компонента слова к другому и патологической персеверации однажды написанного слова. Некоторые дополнительные сведения Вы найдете в главе 7 «Высшие функции нервной системы».
Регистрация активности отдельных нейронов показала, что нейроны премоторной коры низших обезьян активируются до выполнения движения, что может указывать на их связь с программированием этого движения. При этом в структуре нейронной реакции находит отражение направление, в котором будет производиться данное движение (в ответ, например, на условный сигнал). Изучение корреляции нейронных характеристик с параметрами движений рук у обезьяны приводит исследователей к мысли, что в премоторных нейронных сетях содержится своеобразная «библиотека» отдельных целенаправленных движений. Например, если обезьяна подносит рукой ко рту кусочек пищи, то рисунок нейронного разряда не зависит от траектории этого движения (например, из разных точек экстраперсонального пространства). Предполагают, что премоторная кора у приматов участвует в программировании целенаправленных движений (рук) в экстраперсональном пространстве (в пределах вытянутой руки).
Участие в двигательном контроле теменной коры. По современным данным, поле 5 теменной коры также принимает участие как в сенсорных, так и в моторных процессах. Поле 5 часто рассматривают как верхний этаж обработки кинестетической информации. Показано, что при локальном обратимом выключении (например, охлаждением) поля 5 у обезьян нарушались двигательные условные рефлексы, выполнение которых существенно зависело от сохранности проприоцептивного контроля, а при выключении поля 7 страдала также и зрительная обратная связь. У кошек глубокий распад двигательных условных рефлексов на зрительные сигналы происходил только при совместном разрушении полей 5 и 7, тогда как при изолированном разрушении каждого из теменных полей двигательные условные рефлексы заметно не страдали.
Данные, полученные при исследовании нейронных сетей, указывают, что нейроны моторной коры кодируют движение в координатах динамики движения и имеют дирекциональную (их активность определяется направлением движения конечности) настройку. В отличие от этого нейронные сети поля 5 кодируют пространственную траекторию движения, и эти нейроны лишь незначительно модулируют свою активность при воздействии нагрузок на движущуюся руку. Отсюда был сделан вывод, что нейронные сети моторной коры кодируют движение в координатах его динамики, тогда как нейроны теменной коры кодируют пространственную траекторию движения.
Кортико-руброспинальная система двигательного контроля. От красного ядра, которое залегает в толще среднего мозга, берет начало руброспинальный путь. Моторная область коры, откуда берет начало пирамидный тракт, проецируется ипсилатерально на задние 2/3 красного ядра. Сенсомоторная кора (у кошки задняя часть сигмовидной извилины) проецируется ипсилатерально на переднюю область мелкоклеточной части красного ядра,
В ряду позвоночных животных структура красного ядра претерпевает существенные изменения (рис. 5.24). У рептилий и птиц оно однородно и содержит только крупные нейроны. У млекопитающих в дополнение к крупноклеточной (диаметр сомы 50-90 мкм) каудальной части ядра появляется ростральная часть, которая состоит преимущественно из мелких и средних нейронов (диаметр сомы 20-40 мкм). Соотношение этих двух разделов ядра варьирует, но у приматов увеличивается мелкоклеточная часть. Главный эфферентный путь красного ядра — руброспинальный тракт. После перекреста на уровне среднего мозга он спускается в мозговом стволе. В спинном мозге руброспинальный тракт идет в составе латеральных столбов (ростральнее латерального пирамидного тракта) и прослеживается до крестцовых сегментов спинного мозга. Основная часть афферентных проекций на мелкоклеточную часть красного ядра берет начало в зубчатом ядре мозжечка, а на мелкоклеточную часть — в промежуточных ядрах (круглом и пробковидном) мозжечка. Μелкоклеточное подразделение красного ядра
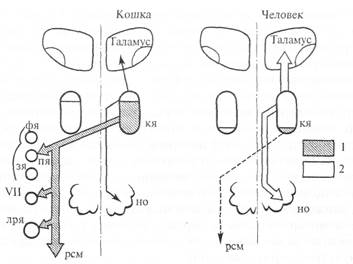
Рис. 5.24. Схема руброспинального тракта.
кя — красное ядро; фя — ядро шатра; пя — промежуточное ядро; зя — зубчатое ядро; VII — ядро лицевого нерва; лря — латеральное ретикулярное ядро; но — нижняя олива; рем — руброспинальный тракт; 1 — крупноклеточная часть кя; 2 — мелкоклеточная часть кя.
образует восходящие проекции на вентролатеральное ядро таламуса, а нисходящие — на нижнюю оливу в продолговатом мозге. Увеличение размеров мелкоклеточной части и уменьшение крупноклеточной части красного ядра происходит на поздних стадиях эволюции и совпадает с появлением неомозжечковых структур и основной оливы. Регресс крупноклеточной части наиболее сильно выражен у человека и совпадает с неразвитостью руброспинального тракта.
Рассечение ствола мозга у четвероногих (кошка, собака) на уровне ростральнее переднего двухолмия не вызывает больших изменений в мышечном тонусе. Однако если разрез проходит на интраколликулярном уровне (изолируется красное ядро), это приводит у четвероногих животных к ригидности (так называемая децеребрационная ригидность). Она развивается по гамма-типу, т. е. обусловлена повышенной импульсацией в гамма-мотонейронах. В отличие от ригидности по альфа-типу эта ригидность пропадает после перерезки дорсальных корешков спинного мозга. Был сделан вывод, что крупноклеточная часть красного ядра осуществляет тормозное влияние на экстензоры постуральных мышц.
Опыты с электрической стимуляцией красного ядра показали, что преимущественно возбуждаются флексоры ипсилатеральной конечности. При стимуляции отдельных точек красного ядра прослеживается соматотопия.
В настоящее время есть основание предполагать, что обе системы двигательного контроля — кортико- и руброспинальные — организованы на уровне нейронного аппарата сегментарных механизмов спинного мозга в значительной степени сходно. Волокна обеих систем моносинаптически возбуждают в основном специализированные промежуточные нейроны латеральных участков серого вещества основания дорсального рога и нейроны промежуточного ядра и уже через эти переключения возбуждают мотонейроны.
Нейрофизиология ретикулоспинальных путей. Начало ретикулоспинальным путям дают нейроны на всем протяжении ретикулярной формации мозгового ствола. Эти пути спускаются в составе латеральных канатиков спинного мозга. Часть волокон в стволе мозга перекрещивается. Волокна ретикулоспинальной системы имеют различный диаметр и обеспечивают проведение возбуждения в широком диапазоне скоростей.
Облегчающие ретикулоспинальные влияния. Американские нейрофизиологи Р. Райнс и Г. Мэгун (1946) установили, что слабое электрическое раздражение ретикулярной формации мозгового ствола и более ростральных отделов мозга усиливает двигательные реакции на раздражение моторной области коры и заметно облегчает коленный рефлекс (рис. 5.25). Эти эффекты опосредованы ретикулоспинальными волокнами, преимущественно идущими в составе латерального канатика. Область, из которой вызываются двусторонние облегчающие влияния, включает значительную часть ретикулярной формации продолговатого мозга, покрышку моста и среднего мозга, центральное серое вещество, субталамус и гипоталамус, а также срединное и интраламинарные ядра таламуса. Облегчающая ретикулоспинальная система играет важную роль в развитии децеребрационной ригидности и спастичности. На децеребрированных кошках (перерезка мозгового ствола на межколликулярном уровне) было показано, что стимуляция участков облегчающей системы ниже уровня перерезки усиливает и без того увеличенные в таких условиях проприоцептивные рефлексы. Этот эффект развивается билатерально. Влияние облегчающей системы мозгового ствола на экстензорный тонус было убедительно показано на кошках с хронической спастичностью, которая развивалась
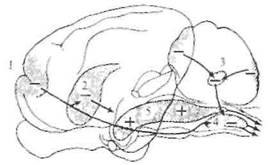
Рис. 5.25. Схема областей мозга, электрическая стимуляция которых вызывает облегчение или торможение двигательной активности.
1 — моторная кора; 2 — полосатые тела; 3— мозжечок; 4, 5 — ретикулярная формация.
в результате удаления двигательной области коры больших полушарий. Спастическое состояние, устраняющееся во время эксперимента на фоне наркоза, не только восстанавливалось, но существенно усиливалось при стимуляции облегчающей системы мозгового ствола вплоть до развития судорог.
Тормозные влияния ретикулярной формации продолговатого мозга.
Значительный вклад в развитие представлений о нейрофизиологических механизмах контроля тонуса внесло открытие Г. Мэгуном и Р. Райнсом (1947) области вентральной ретикулярной формации продолговатого мозга, оказывающей тормозное влияние на разные виды двигательной активности: флексорный, коленный и другие рефлексы. Движения, вызванные раздражением моторной коры, также сильно тормозились. У децеребрированной кошки раздражение указанной области приводило к полной потере тонуса в ригидно разогнутых конечностях. Все тормозные эффекты проявлялись билатерально, но на ипсилатеральной стороне они имели меньший порог. Тормозные эффекты, вызываемые на спинальном уровне, были опосредованы ретикулоспинальными путями, проходившими в переднелатеральной области белого вещества спинного мозга. Активность тормозного механизма продолговатого мозга зависела от высших отделов мозга. При устранении этих влияний, например в результате высокой децеребрации и удаления мозжечка, ретикулоспинальный тормозный механизм переставал оказывать влияние на спинальную активность.
Электрическое раздражение тормозного центра мозгового ствола вызывает в мотонейронах сгибателей и разгибателей торможение. Нисходящие волокна, по которым осуществляется это влияние, проходят в вентральном квадранте спинного мозга в составе вентрального ретикулоспинального пути. Латентное время наблюдаемых эффектов указывает на их полисинаптичекую природу.
Другая группа тормозных эффектов опосредована дорсальной ретикулоспинальной системой, берущей начало примерно в тех же областях мозгового ствола, что и вентральный ретикулоспинальный тракт. После перерезки вентрального квадранта спинного мозга (для устранения влияний вентрального ретикулоспинального пути) стимуляция тормозного центра мозгового ствола уже не вызывает торможение в мотонейронах. По-видимому, непрерывный путь волокон в этой области вообще отсутствует, и тормозные эффекты осуществляются через проприоспинальные (собственные) волокна спинного мозга. Поэтому указанный путь обозначается как дорсальная ретикулоспинальная система.
Таким образом, наиболее древняя система надсегментарного моторного контроля — ретикулярная формация мозгового ствола — через ретикулоспинальные пути оказывает как облегчающие, так и тормозные влияния на весь нейронный аппарат спинного мозга. Наряду с быстропроводящими компонентами этих путей, обеспечивающими прямое действие ретикулярной формации на мотонейроны, волокна с меньшей скоростью проведения оказывают влияния на интернейронную сеть. Ретикулоспинальные пути иннервируют мотонейроны сгибательных и разгибательных мышц. Однако эти влияния оказываются преимущественно на проксимальные мышцы конечностей и мышцы туловища.
Вопросы
1. Кортикоспинальный тракт и его нейрофизиология; эффекты повреждения пирамидного тракта.
2. Нейрофизиология руброспинальной системы.
3. Ретикулоспинальные системы двигательного контроля.
Литература
Костюк П. Г. Структура и функция нисходящих путей спинного мозга. Л.: Наука, 1974.
Мэгун Г, Бодрствующий мозг. М. Мир, 1965
Дата добавления: 2015-01-29; просмотров: 1474;