Принципы координации рефлекторной деятельности
Процесс распространения возбуждения по ЦНС зависит не только от свойств нервных центров, но и от взаимодействия различных центров в процессе реализации как любого рефлекторного акта, так и целенаправленного поведения. Правила этого взаимодействия называются принципами координации рефлекторной деятельности, к которым относят следующие: 1 торможение; 2 субординация; 3 дивергенция; 4 конвергенция; 5 иррадиация возбуждения; 6 концентрация возбуждения; 7 реципрокная иннервация; 8 общий конечный путь; 9 индукция.
1) Торможение – это активный физиологический процесс, который обеспечивается работой нейронов и заключается в подавлении или предотвращении процесса возбуждения. Торможение всегда возникает как следствие процесса возбуждения.
Впервые процесс торможения был продемонстрирован немецкими учеными – братьями Веберами (1845), наблюдавшими резкое замедление работы сердца после раздражения блуждающего нерва. Торможение в ЦНС было открыто русским физиологом И.М. Сеченовым (1863), который увидел резкое увеличение времени рефлекторных реакций после раздражения определенных структур головного мозга. Торможение один из наиболее важных принципов координации рефлекторной деятельности, так как в его отсутствие работа ЦНС, а значит, и всего организма потеряла бы целесообразность. Торможение по-разному проявляется в различных структурах ЦНС, и поэтому выделяют различные его виды.
Торможение всегда связано с взаимодействием нейронов. В зависимости от участия или неучастия специальных тормозных нейронов выделяют: первичное – реализуется с помощью тормозных нейронов; вторичное – реализуется с помощью возбуждающих нейронов.
В зависимости от направления распространения процесса торможения выделяют (рис. 10):
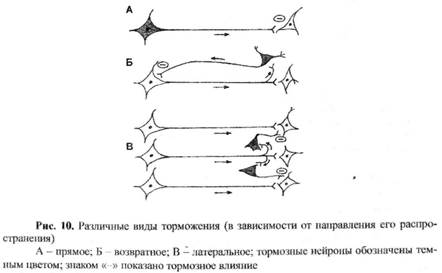 - прямое (афферентное) – торможение поступательно распространяется на последующую структуру (рис.10А);
- прямое (афферентное) – торможение поступательно распространяется на последующую структуру (рис.10А);
- возвратное – с помощью тормозного нейрона торможение «возвращается» на нейрон, вызвавший распространение процесса возбуждения (рис.10Б); имеет большое значение в ограничении процесса возбуждения в различных нервных центрах, препятствует его иррадиации;
- латеральное – оно ограничивает распространение возбуждения в разные стороны от главного направления распространяющегося процесса возбуждения, то есть создает зону торможения вокруг возбуждения (рис. 10В); это торможение играет важную роль в работе анализаторов, ограничивает процесс иррадиации возбуждения и др.;
- реципрокное – это торможение рассмотрено ниже как реципрокная иннервация.
В зависимости от локализации процесса торможение может быть постсинаптическое и пресинаптическое.
Постсинаптическое торможение развивается в результате того, что постсинаптическая мембрана некоторых нейронов теряет свою возбудимость и в связи с этим не может реагировать возбуждением на поступающие к ней импульсы (рис. 11). Этинейроны выключаются из работы, и, тем самым, подавляется или предотвращается распространение возбуждения в ЦНС. Известно два механизма подобного торможения:
А) В основе первого механизма лежит первичное торможение. Среди множества нейронов в ЦНС есть тормозные нейроны. При их возбуждении из пресинаптических терминалей аксонов выделяется медиатор, который не деполяризует, а гиперполяризует постсинаптическую мембрану (возникает ТПСП). Такими медиаторами могут быть бета-глицин, альфа-аланин1 таурин. ГАМК, АХ, дофамин и др. Эти тормозные медиаторы вступают в контакт с теми белками-рецепторами, которые закрывают на постсинаптической мембране калиевые или хлорные каналы. В результате такого контакта эти каналы открываются и ионы К или Сl переходят через мембрану по градиенту концентрации. При этом калий движется по направлению из клетки, а хлор, наоборот, в клетку. Перемещение этих ионов вызывает гиперполяризацию постсинаптической мембраны, а значит, снижение еевозбудимости. Такая мембрана теряет возможность реагировать на импульсы, поступающие по возбуждающим синапсам.
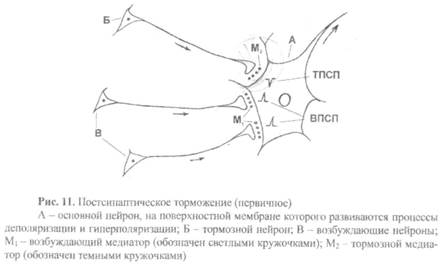 Б) В основе второго механизма лежит вторичное торможение (пессимальное). В этом случае торможение возникает при участии не тормозных, а возбуждающих нейронов. Это возможно только в том случае, когда частота импульсов, проходящих через синапс, очень высока. Механизм вторичного торможения объясняется следующим образом. На постсинаптической мембране в связи с ее длительным возбуждением развивается состояние, подобное катодической депрессии, описанной Б.Ф.Вериго (1883), то есть снижение возбудимости (увеличивается амплитуда мембранного потенциала покоя и порогового потенциала!).
Б) В основе второго механизма лежит вторичное торможение (пессимальное). В этом случае торможение возникает при участии не тормозных, а возбуждающих нейронов. Это возможно только в том случае, когда частота импульсов, проходящих через синапс, очень высока. Механизм вторичного торможения объясняется следующим образом. На постсинаптической мембране в связи с ее длительным возбуждением развивается состояние, подобное катодической депрессии, описанной Б.Ф.Вериго (1883), то есть снижение возбудимости (увеличивается амплитуда мембранного потенциала покоя и порогового потенциала!).
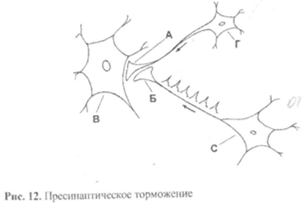 Пресинаптическое торможение заключается в том (рис. 12), что на пресинаптической терминали А обычного возбуждающего синапса (с его помощью может переходить возбуждение от нейрона Г к нейрону В) заканчивается терминаль Б другого нейрона С, образуя на ней аксо-аксональный синапс.
Пресинаптическое торможение заключается в том (рис. 12), что на пресинаптической терминали А обычного возбуждающего синапса (с его помощью может переходить возбуждение от нейрона Г к нейрону В) заканчивается терминаль Б другого нейрона С, образуя на ней аксо-аксональный синапс.
Через этот синапс дополнительный нейрон С оказывает тормозное воздействие на пресинаптический аппарат основного синапса. Это проявляется в снижении возбудимости и проводимости поверхностной мембраны пресинаптического аппарата, а значит, в невозможности ее деполяризации и выделения медиатора в синаптическую щель. В результате этого не происходит передача возбуждения с нейрона Г на нейрон В. Механизм тормозного действия нейрона С на пресинаптичсскую терминаль нейрона Г может быть двояким.
Чаще нейрон С является возбуждающим, но его особенностью является способность к очень высокой импульсной активности (с очень большой частотой – до 1000 импульсов в секунду). В результате на пресинаптической терминами А нейрона Г развивается пессимальное торможение, приводящее к нарушению проведения возбуждения и прекращению выделения медиатора.
Реже нейрон С является обычным тормозным, то есть он выделяет на терминаль А нейрона Г (по механизму постсинаптического торможения) тормозной медиатор, который гиперполяризует поверхностную мембрану и тем снижает ее возбудимость и проводимость, что также приводит к прекращению выделения медиатора.
2) Принцип субординации. В ЦНС много различных центров, участвующих в выполнении какой-либо одной функции. При этом между различными центрами устанавливается определенная согласованность, которая координируется каким-то одним из них. Этому координирующему центру подчиняются другие. Таким образом, существует строгая иерархия между «главными» и «подчиненными» центрами. Например, при непроизвольном дыхании дыхательному центру продолговатого мозга подчиняются мотонейроны спинного мозга, иннервирующие дыхательные мышцы. При произвольном дыхании дыхательный центр на определенное время может подчиняться коре больших полушарий (произвольная задержка дыхания, дыхание при разговоре и др.).
3) Суть принципа реципрокной иннервации в межцентральных взаимоотношениях нейронов. Например, произвольные и непроизвольные движения могут быть реализованы только при условии согласованно работы мышц-антагонистов. Например, при сгибании конечности мышцы-сгибатели должны сокращаться, а мышцы-разгибатели расслабляться. Поэтому при поступлении возбуждения к нервному центру (с рецепторов или из высших отделов ЦНС) один и тот же поток импульсов возбуждает 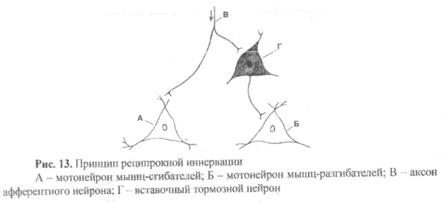 альфа-мотонейроны мышц сгибателей, но одновременно, действуя через вставочные тормозные нейроны, тормозит альфа-мотонейроны мышц разгибателей (рис. 13). Причем на контралатеральной конечности происходят противоположные изменения, так как тот же самый поток импульсов, переходя на противоположную сторону спинного мозга, возбуждает уже альфа-мотонейроны мышц-разгибателей и тормозит альфа-мотонейроны мышц-сгибателей.
альфа-мотонейроны мышц сгибателей, но одновременно, действуя через вставочные тормозные нейроны, тормозит альфа-мотонейроны мышц разгибателей (рис. 13). Причем на контралатеральной конечности происходят противоположные изменения, так как тот же самый поток импульсов, переходя на противоположную сторону спинного мозга, возбуждает уже альфа-мотонейроны мышц-разгибателей и тормозит альфа-мотонейроны мышц-сгибателей.
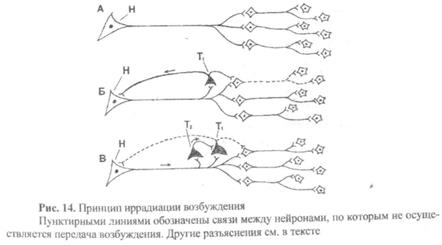 4) Принцип «иррадиация возбуждения». ЦНС состоит из совокупности нейронов, связанных друг с другом благодаря множеству синаптических связей. Эти связи позволяют, вызвав возбуждение только одного нейрона (рис. 14А, Н), добиться возбуждения всей ЦНС. Однако в здоровом организме в норме этого не происходит благодаря наличию тормозных нейронов (рис. 14Б, Т1), включенных в цепь нейронов и оказывающих тормозное влияние на соседние нейроны, ограничивая их возбуждение, а также оказывая тормозное влияние (возвратное торможение) на первоначально возбужденный нейрон (рис. 14Б, Н). Как видим, тормозные нейроны ограничивают не только распространение (иррадиацию) возбуждения, но и его интенсивность.
4) Принцип «иррадиация возбуждения». ЦНС состоит из совокупности нейронов, связанных друг с другом благодаря множеству синаптических связей. Эти связи позволяют, вызвав возбуждение только одного нейрона (рис. 14А, Н), добиться возбуждения всей ЦНС. Однако в здоровом организме в норме этого не происходит благодаря наличию тормозных нейронов (рис. 14Б, Т1), включенных в цепь нейронов и оказывающих тормозное влияние на соседние нейроны, ограничивая их возбуждение, а также оказывая тормозное влияние (возвратное торможение) на первоначально возбужденный нейрон (рис. 14Б, Н). Как видим, тормозные нейроны ограничивают не только распространение (иррадиацию) возбуждения, но и его интенсивность.
Однако при действии сильных раздражителей (рис. 14В) в процессе возбуждения участвует тем большее количество нейронов, чем больше сила действующего раздражителя. Это происходит потому, что в процесс возбуждения вовлекаются менее возбудимые тормозные нейроны (Т2), которые тормозят тормозные нейроны (Т1) и тем самым «снимают» торможение с ранее заторможенных нейронов и способствуют как распространению процесса возбуждении, так и усилению его интенсивности.
Подобный механизм развития иррадиации возбуждения отмечается при отравлении стрихнином, при бешенстве и других состояниях, когда происходит выключение тормозных (Т1) нейронов.
5) Концентрация возбуждения – это процесс, обратный иррадиации возбуждения. Смысл его заключается в том, что в нервном центре, ответственном за регуляцию какой-либо физиологической функции, количество активно функционирующих нейронов с течением времени уменьшается, то есть ту же самую «работу» выполняет меньшее количество нейронов. Например, при выработке условных рефлексов в начальной фазе (генерализация условных рефлексов) преобладают процессы иррадиации возбуждения, вовлекающие в процесс осуществления рефлекторной реакции множество нейронов. Но затем, по мере повторения условно-рефлекторной реакции, когда она становится «заученным» действием, начинают преобладать процессы концентрации возбуждения, так как условно-рефлекторная реакция уже может быть обеспечена меньшим количеством «обученных» нейронов, специализирующихся на анализе только конкретного условного раздражителя. Второстепенные нейроны постепенно выключаются из цепи условно-рефлекторной реакции в связи с возникающим дифференцировочным торможением (обеспечивает отсутствие условно-рефлекторной реакции на раздражители, похожие на условный, но отличающиеся от него но биологическому значению).
6) Принцип дивергенции. Растяжение единственной мышцы приводит к возбуждению нескольких сотен чувствительных нейронов, каждый из которых активирует от 100 до 150 мотонейронов. Этот способ взаимодействия нервных клеток, при котором один нейрон многочисленными ветвями аксона действует на большое количество других нейронов, называется дивергенцией.
7) 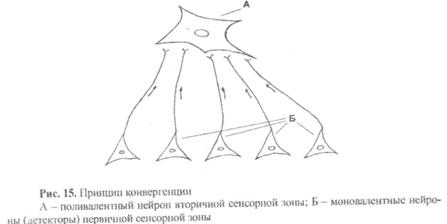 Принцип конвергенции. Здесь под конвергенцией понимают схождение различной афферентной информации в одних и тех же нервных центрах полимодальных или поливалентных нейронах. Это дает возможность мозгу проводить более тонкую аналитико-синтетическую деятельность по оценке действующих на организм раздражителей и выбирать наиболее оптимальный вариант поведения. Так, на рис. 15 показана конвергенция, имеющая место в корковом отделе анализатора. Суть ее заключается в том, что от различных моновалентных нейронов первичной сенсорной зоны (они осуществляют анализ действующих раздражителей) возбуждение поступает (конвергирует) к одному нейрону вторичной сенсорной зоны (здесь идут процессы синтеза ранее проанализированной информации).
Принцип конвергенции. Здесь под конвергенцией понимают схождение различной афферентной информации в одних и тех же нервных центрах полимодальных или поливалентных нейронах. Это дает возможность мозгу проводить более тонкую аналитико-синтетическую деятельность по оценке действующих на организм раздражителей и выбирать наиболее оптимальный вариант поведения. Так, на рис. 15 показана конвергенция, имеющая место в корковом отделе анализатора. Суть ее заключается в том, что от различных моновалентных нейронов первичной сенсорной зоны (они осуществляют анализ действующих раздражителей) возбуждение поступает (конвергирует) к одному нейрону вторичной сенсорной зоны (здесь идут процессы синтеза ранее проанализированной информации).
Различают следующие виды конвергенции:
- мультисенсорная – это конвергенция различной сенсорной информации на одних и тех же подкорковых структурах или участках ассоциативной коры, где происходит взаимодействие в работе различных анализаторов (например, зрительного, слухового, вкусового и т.д.).
 - мультибиологическая – конвергенция информации различной биологической модальности (например, наличие раздражителей, одновременно вызывающих пищедобывательное и оборонительное поведение) на одних и тех же структурах ассоциативной коры.
- мультибиологическая – конвергенция информации различной биологической модальности (например, наличие раздражителей, одновременно вызывающих пищедобывательное и оборонительное поведение) на одних и тех же структурах ассоциативной коры.
- сенсорно-биологическая – это конвергенция информации разной сенсорной и биологической модальности. Например, несмотря на то, что человек голоден, для удовлетворения пищевой потребности необходимы определенные условия.
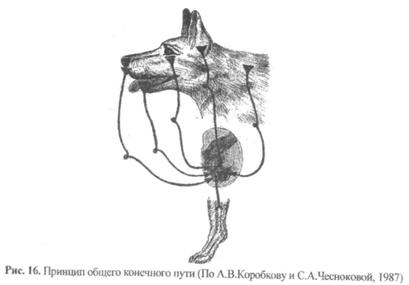
 8) Принцип «общий конечный путь» был сформулирован английским физиологом Ч. Шеррингтоном (1897) как принцип «борьбы за общий конечный путь». Известно, что афферентных входов в ЦНС значительно больше, чем эфферентных выходов, и на одних и тех же моторных нейронах (например, спинного мозга) сходится информация с большого количества разных рецепторов. На тех же мотонейронах заканчиваются эфферентные пути от стволовых и корковых структур головного мозга. Таким образом, моторные нейроны являются общим конечным путем, через который реализуются разные рефлекторные, непроизвольные и произвольные движения (рис. 16).
8) Принцип «общий конечный путь» был сформулирован английским физиологом Ч. Шеррингтоном (1897) как принцип «борьбы за общий конечный путь». Известно, что афферентных входов в ЦНС значительно больше, чем эфферентных выходов, и на одних и тех же моторных нейронах (например, спинного мозга) сходится информация с большого количества разных рецепторов. На тех же мотонейронах заканчиваются эфферентные пути от стволовых и корковых структур головного мозга. Таким образом, моторные нейроны являются общим конечным путем, через который реализуются разные рефлекторные, непроизвольные и произвольные движения (рис. 16).
Например, один и тот же мотонейронный пул (совокупность мотонейронов, иннервирующих одну мышцу) может участвовать в одних случаях в защитной, а в других – в агрессивной реакции, а может и в обычной произвольной двигательной деятельности.
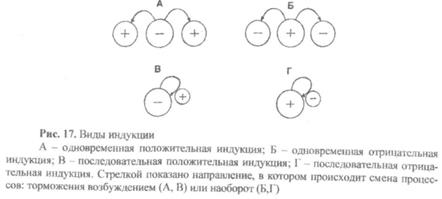
9)Принцип индукции. В работе ЦНС сосуществуют два противоположных по физиологическому смыслу процесса: возбуждение и торможение. Индукция – это провокация (наведение, вызывание) одним процессом (например, возбуждением) другого – противоположного (торможения) (рис. 17).
Индукция может быть положительной и отрицательной. О положительной индукции говорят в том случае, когда процесс торможения провоцирует возникновение процесса возбуждения (рис. 17 А,В). В противоположном случае, когда процесс возбуждения вызывает возникновение процесса торможения, речь идет об отрицательной индукции (рис. 17 Б,Г). Положительная и отрицательная индукция могут быть одновременной и последовательной
Об одновременной индукции можно говорить в том случае, когда в одно и то же время два центра находятся в противоположных состояниях, то есть один из них возбужден, а другой заторможен или наоборот. Одновременная индукция может быть положительной (рис. I7A) и отрицательной (рис. 17Б). О последовательной индукции можно говорить в том случае, когда в одном и том же центре один процесс в течение времени меняется на противоположные. Последовательная индукция может быть положительной (рис. 17В) и отрицательной (рис.17Г).
Дата добавления: 2015-01-21; просмотров: 8333;