Мембранный потенциал покоя
В покое на наружной стороне плазматической мембраны располагается тонкий слой положительных зарядов, а на внутренней стороне – отрицательных. Разность между ними называется мембранным потенциалом покоя.Если считать наружный заряд равным нулю, то разность зарядов между наружной и внутренней поверхностями у большинства нейронов оказывается близкой к -65 мВ, хотя она и может у отдельных клеток варьировать от -40 до -80 мВ.
Возникновение этой разности зарядов обусловлено неодинаковым распределением ионов калия, натрия и хлора внутри клетки и снаружи её, а также большей проницаемостью покоящейся клеточной мембраны лишь для ионов калия.
У возбудимых клеток мембранный потенциал покоя (МПП) способен сильно изменяться и эта способность является основой для возникновения электрических сигналов. Уменьшение мембранного потенциала покоя, например, с -65 до -60 мВ, называется деполяризацией, а увеличение, например, с -65 до -70 мВ, – гиперполяризацией.
 Если деполяризация достигнет некоторого критического уровня, например -55 мВ, то проницаемость мембраны для ионов натрия на короткое время становится максимальной, они устремляются в клетку и в связи с этим трансмембранная разность потенциалов стремительно уменьшается до 0, а затем приобретает положительное значение. Это обстоятельство приводит к закрытию натриевых каналов и стремительному выходу из клетки ионов калия через предназначенные только для них каналы: в результате восстанавливается первоначальная величина мембранного потенциала покоя. Эти быстро происходящие изменения мембранного потенциала покоя называются потенциалом действия. Потенциал действия является приводящимся электрическим сигналом, он быстро распространяется по мембране аксона до самого его окончания, причём нигде не меняет свою амплитуду.
Если деполяризация достигнет некоторого критического уровня, например -55 мВ, то проницаемость мембраны для ионов натрия на короткое время становится максимальной, они устремляются в клетку и в связи с этим трансмембранная разность потенциалов стремительно уменьшается до 0, а затем приобретает положительное значение. Это обстоятельство приводит к закрытию натриевых каналов и стремительному выходу из клетки ионов калия через предназначенные только для них каналы: в результате восстанавливается первоначальная величина мембранного потенциала покоя. Эти быстро происходящие изменения мембранного потенциала покоя называются потенциалом действия. Потенциал действия является приводящимся электрическим сигналом, он быстро распространяется по мембране аксона до самого его окончания, причём нигде не меняет свою амплитуду.
Кроме потенциалов действия в нервной клетке, вследствие изменения её мембранной проницаемости, могут возникать местные или локальные сигналы: рецепторный потенциал и постсинаптический потенциал. Их амплитуда значительно меньше, чем у потенциала действия, кроме того, она существенно уменьшается при распространении сигнала. По этой причине местные потенциалы и не могут распространяться по мембране далеко от места своего возникновения.
Работой натрий-калиевого насоса в клетке создаётся высокая концентрация ионов калия, а в клеточной мембране для этих ионов есть открытые каналы. Выходящие из клетки по концентрационному градиенту ионы калия увеличивают количество положительных зарядов на наружной поверхности мембраны. В клетке много крупномолекулярных органических анионов и потому изнутри мембрана оказывается заряженной отрицательно. Все остальные ионы могут проходить через покоящуюся мембрану в очень небольшом количестве, их каналы, в основном, закрыты. Следовательно, потенциал покоя обязан своим происхождением, главным образом, току ионов калия из клетки.
| |
Электрические сигналы: входной, объединённый, проводящийся и выходной
 Нейроны вступают в контакты с определёнными клетками-мишенями, причём цитоплазма контактирующих клеток не соединяется и между ними всегда сохраняется синаптическая щель.
Нейроны вступают в контакты с определёнными клетками-мишенями, причём цитоплазма контактирующих клеток не соединяется и между ними всегда сохраняется синаптическая щель.
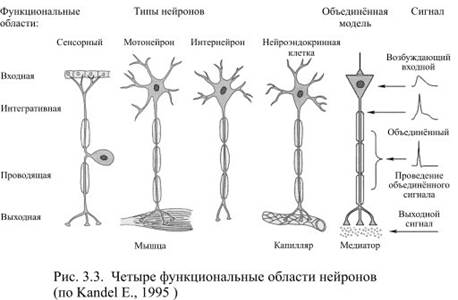
Современный вариант нейронной теории связывает определённые части нервной клетки с характером возникающих в них электрических сигналов. В типичном нейроне есть четыре определяемые морфологически области: дендриты, сома, аксон и пресинаптическое окончание аксона. При возбуждении нейрона в нём последовательно появляется четыре разновидности электрических сигналов: входной, объединённый, проводящийся и выходной (рис. 3.3). Каждый из этих сигналов возникает только в определённой морфологической области.
Входными сигналами являются либо рецепторный, либо постсинаптический потенциал. Рецепторный потенциал образуется в окончаниях чувствительного нейрона, когда на них действует определённый стимул: растяжение, давление, свет, химическое вещество и т.п. Действие стимула вызывает открытие определённых ионных каналов мембраны, а последующий ток ионов через эти каналы изменяет первоначальное значение мембранного потенциала покоя; в большинстве случаев происходит деполяризация. Эта деполяризация и является рецепторным потенциалом, её амплитуда пропорциональна силе действующего стимула.
Рецепторный потенциал может распространяться от места действия стимула вдоль мембраны на относительно небольшое расстояние – амплитуда рецепторного потенциала уменьшается по мере удаления от места действия стимула, а затем деполяризующий сдвиг и вовсе исчезнет.
Вторая разновидность входного сигнала – постсинаптический потенциал. Он образуется на постсинаптической клетке после того, как возбуждённая пресинаптическая клетка отправит для неё нейромедиатор. Добравшись путём диффузии до постсинаптической клетки, медиатор присоединяется к специфическим белкам-рецепторам её мембраны, что вызывает открытие ионных каналов. Возникший в связи с этим ток ионов через постсинаптическую мембрану изменяет первоначальное значение мембранного потенциала покоя – этот сдвиг и является постсинаптическим потенциалом.
В одних синапсах такой сдвиг представляет собой деполяризацию и, если она достигнет критического уровня, то постсинаптический нейрон возбуждается. В других синапсах возникает противоположный по направленности сдвиг: постсинаптическая мембрана гиперполяризуется: величина мембранного потенциала становится больше и уменьшить её до критического уровня деполяризации становится труднее. Такую клетку трудно возбудить, она заторможена. Таким образом, деполяризующий постсинаптический потенциал является возбуждающим, а гиперполяризующий – тормозным. Соответственно этому и сами синапсы подразделяются на возбуждающие (вызывающие деполяризацию) и тормозные (вызывающие гиперполяризацию).
Вне зависимости от того, что происходит на постсинаптической мембране: деполяризация или гиперполяризация, величина постсинаптических потенциалов всегда пропорциональна количеству подействовавших молекул медиатора, но обычно их амплитуда невелика. Так же, как и рецепторный потенциал, они распространяются вдоль мембраны на очень небольшое расстояние, т.е. тоже относятся к местным потенциалам.
Таким образом, входные сигналы представлены двумя разновидностями местных потенциалов, рецепторным и постсинаптическим, а возникают эти потенциалы в строго определённых областях нейрона: либо в чувствительных окончаниях, либо в синапсах. Чувствительные окончания принадлежат сенсорным нейронам, где рецепторный потенциал возникает под действием внешних раздражителей. Для интернейронов, а также для эфферентных нейронов входным сигналом может быть только постсинаптический потенциал.
Объединённый сигнал может возникнуть только в таком участке мембраны, где достаточно много ионных каналов для натрия. В этом отношении идеальным объектом является аксонный холмик – место отхождения аксона от тела клетки, поскольку именно здесь самая высокая во всей мембране плотность каналов для натрия. Такие каналы являются потенциалзависимыми, т.е. открываются лишь тогда, когда исходное значение потенциала покоя достигнет критического уровня. Типичное для среднестатистического нейрона значение потенциала покоя составляет приблизительно -65 мВ, а критический уровень деполяризации соответствует примерно -55 мВ. Стало быть, если удастся деполяризовать мембрану аксонного холмика с -65 мВ до -55 мВ, то там возникнет потенциал действия.
Деполяризовать мембрану способны входные сигналы, т.е. либо постсинаптические потенциалы, либо рецепторные. В случае рецепторных потенциалов местом возникновения объединённого сигнала является ближайший к чувствительным окончаниям перехват Ранвье, где наиболее вероятна деполяризация до критического уровня. Каждый чувствительный нейрон имеет множество окончаний, являющихся ветвями одного отростка. И, если в каждом из этих окончаний при действии стимула возникает очень небольшой по амплитуде рецепторный потенциал и распространяется к перехвату Ранвье с уменьшением амплитуды, то он является лишь малой частью общего деполяризующего сдвига. От каждого чувствительного окончания в одно и то же время перемещаются к ближайшему перехвату Ранвье эти небольшие рецепторные потенциалы, а в области перехвата все они суммируются. Если общая сумма деполяризующего сдвига будет достаточной, то в перехвате возникнет потенциал действия.
Постсинаптические потенциалы, возникающие на дендритах, так же невелики, как и рецепторные потенциалы и так же уменьшаются при распространении от синапса до аксонного холмика, где может возникнуть потенциал действия. Кроме того, на пути распространения постсинаптических потенциалов по телу клетки могут оказаться тормозные гиперполяризующие синапсы и потому возможность деполяризации мембраны аксонного холмика на 10 мВ кажется маловероятной. Тем не менее, этот результат регулярно достигается в результате суммации множества небольших постсинаптических потенциалов, возникающих одновременно в многочисленных синапсах, образованных дендритами нейрона с окончаниями аксонов пресинаптических клеток.
Таким образом, объединённый сигнал возникает, как правило, вследствие суммации одновременно образовавшихся многочисленных местных потенциалов. Такая суммация происходит в том месте, где особенно много потенциалзависимых каналов и поэтому легче достигается критический уровень деполяризации. В случае интеграции постсинаптических потенциалов таким местом является аксонный холмик, а суммация рецепторных потенциалов происходит в ближайшем от чувствительных окончаний перехвате Ранвье (или близко расположенным к ним участком немиелинизированного аксона). Область возникновения объединённого сигнала называется интегративной или триггерной.
Накопление небольших деполяризующих сдвигов молниеносно трансформируется в интегративной зоне в потенциал действия, который является максимальным электрическим потенциалом клетки и возникает по принципу "всё или ничего". Это правило надо понимать так, что деполяризация ниже критического уровня не приносит никакого результата, а при достижении этого уровня всегда, независимо от силы стимулов, обнаруживается максимальный ответ: третьего не дано.
Проведение потенциала действия. Амплитуда входных сигналов пропорциональна силе подействовавшего стимула или количеству выделившегося в синапсе нейромедиатора – такие сигналы называют градуальными. Их длительность определяется длительностью стимула или присутствия медиатора в синаптической щели. Амплитуда и длительность потенциала действия от этих факторов не зависят: оба этих параметра всецело определяются свойствами самой клетки. Стало быть, любая комбинация входных сигналов, любой вариант суммации, при единственном условии деполяризации мембраны до критического значения, вызывает один и тот же стандартный образец потенциала действия в триггерной зоне. Он всегда имеет максимальную для данной клетки амплитуду и примерно одинаковую длительность, сколько бы раз ни повторялись вызывающие его условия.
Возникнув в интегративной зоне, потенциал действия быстро распространяется по мембране аксона. Это происходит благодаря появлению локального электрического тока. Поскольку деполяризованный участок мембраны оказывается иначе заряженным, чем соседствующий с ним, между полярно заряженными участками мембраны возникает электрический ток. Под действием этого локального тока деполяризуется до критического уровня соседний участок, что вызывает появление потенциала действия и в нём. В случае миелинизированного аксона таким соседним участком мембраны является ближайший к триггерной зоне перехват Ранвье, затем следующий, и потенциал действия начинает "перепрыгивать" от одного перехвата к другому со скоростью, достигающей 100 м/с.
Разные нейроны могут многим отличаться друг от друга, но возникающие в них потенциалы действия различить очень трудно, чаще невозможно. Это в высшей степени стереотипный сигнал у самых разных клеток: сенсорных, интернейронов, моторных. Эта стереотипия свидетельствует о том, что сам потенциал действия не содержит никаких сведений о природе породившего его стимула. О силе стимула свидетельствует частота возникающих потенциалов действия, а определением природы стимула занимаются специфические рецепторы и хорошо упорядоченные межнейронные связи.
Таким образом, возникший в триггерной зоне потенциал действия быстро распространяется по ходу аксона к его окончанию. Это передвижение связано с образованием локальных электрических токов, под влиянием которых потенциал действия как бы заново возникает в соседнем участке аксона. Параметры потенциала действия при проведении по аксону нисколько не меняются, что позволяет передавать информацию без искажений. Если аксоны нескольких нейронов оказываются в общем пучке волокон, то по каждому из них возбуждение распространяется изолированно.
Выходной сигнал адресуется другой клетке или одновременно нескольким клеткам и в подавляющем большинстве случаев представляет собой выделение химического посредника – медиатора. В пресинаптических окончаниях аксона заранее запасённый медиатор хранится в синаптических пузырьках, которые накапливаются в специальных участках – активных зонах. Когда потенциал действия добирается до пресинаптического окончания, содержимое синаптических пузырьков путём экзоцитоза опорожняется в синаптическую щель.
Химическими посредниками передачи информации могут служить разные вещества: небольшие молекулы, как, например, ацетилхолин или глутамат, либо достаточно крупные молекулы пептидов – все они специально синтезируются в нейроне для передачи сигнала. Попав в синаптическую щель, медиатор диффундирует к постсинаптической мембране и присоединяется к её рецепторам. В результате связи рецепторов с медиатором изменяется ионный ток через каналы постсинаптической мембраны, а это приводит к изменению значения потенциала покоя постсинаптической клетки, т.е. в ней возникает входной сигнал – в данном случае постсинаптический потенциал.
Таким образом, почти в каждом нейроне, независимо от его величины, формы и занимаемой в цепи нейронов позиции, можно обнаружить четыре функциональные области: локальную рецептивную зону, интегративную, зону проведения сигнала и выходную или секреторную зону (рис. 3.3).
Дата добавления: 2015-01-21; просмотров: 3831;