МЕДИАТОРЫ ДЕЙСТВИЯ ГОРМОНОВ
ОБЩИЙ ОЧЕРК: ГИПОТЕЗА «ВТОРОГО МЕДИАТОРА»
Перенос наружных (т. е. внеклеточных) гормональных «сигналов» во внутреннюю среду клетки после лигандрецепторного взаимодействия осуществляется «вторыми медиаторами», которые образуются на мембранном уровне. Первоначальная формулировка этой концепции, принадлежащая Sutherland, показана на рис. 4—2, где представлен общий механизм, с помощью которого пептидные гормоны, действующие на клеточную поверхность, стимулируют образование «вторых медиаторов», таких, как цАМФ. Вначале эта общая схема действия пептидных гормонов на мембранные рецепторы и аденилатциклазу была предложена на основании исследований, проведенных с катехоламинами, вазопрессином, глюкагоном и АКТГ, но затем было показано, что многие пептидные гормоны вызывают активацию аденилатциклазы и повышение внутриклеточного уровня цАМФ. Однако для некоторых пептидных гормонов, таких, как ангиотензин, инсулин и СТГ, механизм активации клеток-мишеней остается невыясненным (табл. 4—1). Даже в тех тканях, в которых «вторым медиатором» действия пептидных гормонов принято считать цАМФ, неполностью ясен способ, которым одна эффекторная система транслируется в целый спектр реакций отдельных клеток-мишеней. Этот вопрос осложняется двойственной природой многих пептидных и белковых гормонов, которые, с одной стороны, являются трофическими факторами с их поздними влияниями на рост и дифференцировку клеток, а с другой, выступают в роли быстрых регуляторов таких клеточных реакций, как стероидогенез и секреция гормонов. Схема этих медленных и быстрых эффектов гормонов, оказывающих как поздние ростовые, так и кратковременные метаболические влияния [27], приведена на рис. 4—9. Связь между двумя указанными аспектами действия пептидных гормонов выяснена недостаточно полно, и большинство наших знаний о клеточных реакциях на гормонрецепторное взаимодействие основано на результатах исследования острых эффектов пептидных гормонов на их клетки-мишени.
Внутриклеточные сигнальные молекулы, генерируемые гормон-рецепторным взаимодействием, действуют затем на мембранные и цитоплазматические белки, изменяя скоростьограничивающие этапы эффекторных систем, контролирующих реакции клетки-мишени. Лучше всего охарактеризованными медиаторами являются цАМФ и кальций, выступающие в роли внутриклеточных трансмиттеров и приводящие метаболизм клетки-мишени в соответствии с потребностями в реакции на внешнюю стимуляцию такими факторами, как гормоны и другие лиганды. Если кальциевый стимул обеспечивается повышением концентрации кальция в цитозоле за счет поглощения кальция из внеклеточной жидкости или перераспределения внутриклеточного кальция, то циклические нуклеотиды образуются из АТФ и ГТФ за счет активации циклазных ферментов, расположенных в плазматической мембране и цитозоле соответственно. Таким образом, образование цАМФ непосредственно связано с насыщением рецепторов клеточной поверхности и активацией аденилатциклазы, локализующейся на внутренней поверхности липидного бислоя плазматической мембраны. В отличие от этого, гуанилатциклаза расположена преимущественно в цитозоле и не активируется непосредственно гормональными «стимулами». Кальций оказывает важное действие как на гуанилатциклазную активность, так и на разрушение гЦМФ фосфодиэстеразой, и внутриклеточная концентрация кальция служит, вероятно, основным регулятором уровня цАМФ и цГМФ в клетке [28].
Таблица 4—1. Пептидные гормоны и аденилатциклаза
1. Гормоны, действующие через цАМФ как «второй медиатор» а. Адренокортикотропный гормон (АКТГ, кортикотропин) б. Лютеинизирующий гормон (ЛГ, лютеотропин) в. Хорионический гонадотропин человека (ХГЧ) г. Фолликулостимулирующий гормон (ФСГ, фоллитропин) д. Гонадотропин сыворотки жеребой кобылы (ГСЖК) е. Тиротропный гормон (ТТГ, тиротропин) ж. Липотропин (ЛПГ) з. Меланоцитстимулирующий гормон (МСГ, меланотропин) и. Антидиуретический гормон (АДГ, вазопрессин) к. Тиротропин-рилизинг гормон (ТРГ) л. Паратиреоидный гормон (ПТГ) м. Кальцитонин (КТ) н. Глюкагон о. Катехоламины (b-адренергические эффекты) п. Простагландины (ПГЕ1)
2. Гормоны, действующие, вероятно, не через цАМФ а. Ангиотензин II б. Гонадотропин-рилизинг гормон (ГнРГ, рилизинг-гормон лютеинизирующего гормона, РГ-ЛГ)
3. Гормоны, действующие не через цАМФ а. Инсулин б. Инсулиноподобные факторы роста в. Соматомедин г. Фактор роста эпидермиса (ФРЭ) д. Фактор роста фибробластов (ФРФ) е. Активность, стимулирующая размножение (АСР) ж. Фактор роста нервов (ФРН) з. Гормон роста (СТГ) и. Пролактин к. Плацентарный лактоген л. Окситоцин м. Соматостатин н. Катехоламины (a-адренергические эффекты) о. Простагландины (ПГF2a) [3]
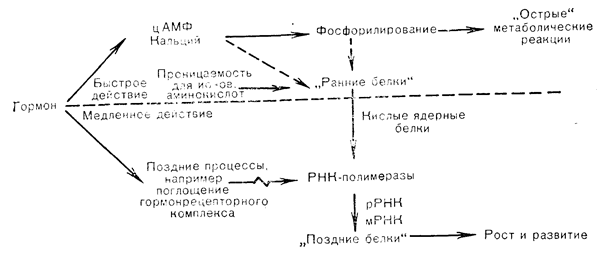
Рис. 4—9. Общая схема интеграции ранних и поздних реакций на действие гормонов, вызывающих как острые метаболические сдвиги, так и долговременные изменения роста и развития (Tata [27] в модификации).
Роль кальция в сопряжении стимула и реакции при активации сократительных и секреторных процессов была установлена достаточно надежно. Впоследствии, однако, выяснение широко распространенных эффектов цАМФ в качестве «второго медиатора» обусловило тенденцию к умалению роли кальция как внутриклеточно го регулятора. Позднее было показано значение взаимодействий между циклическими нуклеотидами и кальцием [29], и вновь стали подчеркивать примат этого иона в качестве медиатора активации многих процессов в клетках-мишенях [30].
Дата добавления: 2015-01-19; просмотров: 2326;