Біофізика м'язового скорочення
Життєдіяльність людини, тварин, птахів|птиць|, риб, рослин і інших біологічних об'єктів нерозривно пов'язана з механічними рухами (переміщеннями). Всі види руху у біооб'єктах можна розділити на дві групи: макрорухи і мікрорухи. До макрорухів слід віднести: різні види рухів тіла, кровообіг, дихання. До мікрорухів відносять транспорт речовин через мембрани, коливальні, циркуляційні і фонтануючі рухи цитоплазми, водно-електролітний обмін у капілярних судинах|посудинах| і інші мікропроцеси у клітинах|клітинах|, тканинах та органах. Макрорухи здійснюються завдяки наявності м'язових утворень (м'язів), що володіють спеціальним білковим скоротливим|скорочувальним| комплексом — актоміозином|.
На походження мікрорухів дотепер|до цих пір| немає єдиної точки зору, проте|однак| найбільш вірогідною вважається|лічиться| гіпотеза функціювання скоротливих|скорочувальних| елементів, подібних до м'язових.
У|із| приведеному короткому огляді показана величезна роль вчення про механізм роботи м'язів, як однієї з основних форм існування живої|жвавої| матерії. Знання суті|сутності,єства| м'язового скорочення є фундаментом вчення про пряме перетворення хімічної енергії в механічну, без проміжного перетворення її у теплову або електричну енергію. Це явище представляє|уявляє| великий технічний інтерес, відкриваючи|відчиняючи| перспективи створення|створіння| нового типу двигунів з|із| достатньо|досить| великим коефіцієнтом корисної дії.
Скелетний м'яз хребетних|хребцевих| складається з декількох тисяч паралельних м'язових волокон діаметром (10-100 мкм|), пов’язнених в загальну|спільну| оболонку. Волокна є|з'являються,являються| абсолютно пружними елементами і сприймають напругу|напруження| при скороченні м'яза. На кожному кінці м'яза вони переходять у сухожилля, що закріплені на кістковій тканині. Через кінцеву пластинку|платівку| кожного м'язового волокна приєднуються закінчення нервових волокон. Нервовий імпульс від ЦНС| приводить|призводить,наводить| до роздратування групи м'язових волокон (рухових одиниць).
М'язове волокно, у свою чергу|своєю чергою|, містить|утримує| від 1000 до 2000 паралельних ниток (міофібрил) діаметром близько 1 мкм|. Весь пучок міофібрил розміщено|ув'язнений| у мембрані — плазмолемі. Плазмолема, подібно до мембран інших клітин|клітин|, складається з трьох шарів білків та ліпідів, загальною|спільною| товщиною близько 10 нм| і електрично поляризована. Зверху плазмолема покрита тонким шаром колагенових ниток, що володіють пружними властивостями.
Окрім|крім| міофібрил у м'язовому волокні міститься|утримується| багато ядер, розташованих|схильних| поблизу плазмолеми, і велика кількість мітохондрій, що знаходяться|перебувають| між міофібрилами. Ядра складаються з трубочок і бульбашок (цитоплазматичний| ретикулум|), які виділяють кальцій при збудженні м'язового волокна і видаляють|знищують,віддаляють| його з|із| саркоплазми при припиненні м'язового збудження. Мітохондрії є|з'являються,являються| центральними утвореннями макроергічних з'єднань|сполучень,сполук|, перш за все|передусім| АТФ. Молекули АТФ поступають|надходять| із|із| мітохондрій через саркоплазму до міофібрил у момент скорочення.
Основною надмолекулярною руховою структурою м'язових волокон є|з'являється,являється| саркомер. Саркомери розташовані|схильні| у м'язовому волокні послідовно один за іншим. Структура саркомера| показана на мал. 6.19.
Кожен саркомер (1) обмежений Z — пластинками|платівками|. Між ними знаходяться|перебувають| тонкі нитки, складені з|із| молекул білка F-актина, і товсті нитки, утворені молекулами білка міозину.
Нитки мають гексагональну упаковку (2), так, що кожна товста нитка взаємодіє з|із| 6-ма тонкими нитками, а кожна тонка нитка взаємодіє з|із| 3-мя| товстими нитками. Тонка нитка має структуру подвійної суперспіралі (3), а кожна спіраль складається з глобули| білка G-актина.

Малюнок 6.19-Структура саркомера
Глобула — це просторове розташування полімерної нитки, у якій окремі ланки мають певну просторову структуру і розташування. Товсті нитки (4) також на 50 % складаються з спіралі молекул міозину. Проте|однак|, в товстій нитці є, так звані, «шарнірні» ділянки, де у суперспіраль вбудовані глобулярні головки|голівки|, розташовані|схильні| радіально до вісі товстої нитки. Ці головки|голівки| розташовані|схильні| по обидві сторони товстої нитки (їх називають містками). Два суміжних містки розташовуються під кутом 120° один до одного; отже ділянка з|із| 4 містків є|з'являється,являється| елементом, який повторюється, і має довжину 43 нм|.
Скорочення (укорочення) м'язового волокна відбувається|походить| за рахунок зменшення довжини саркомерів по наступній|такій| схемі. При роздратуванні м'язового волокна нервовим імпульсом з|із| саркоплазматичного ретикулума| поступають|надходять| іони кальцію, які активізують певні мікроділянки тонких ниток. Як результат – актинові| центри стають доступними для взаємодії з|із| міозиновими| містками.
Структура міозинової| нитки не змінюється, а міозинові| містки змінюють|зраджують| свій напрям|направлення| (нахиляються|схиляються| вліво) і проштовхують товсту нитку ближче до Z — пластинки|платівці|, збуджена ж область тонких ниток зміщується на один крок вправо (мал. 6.20).
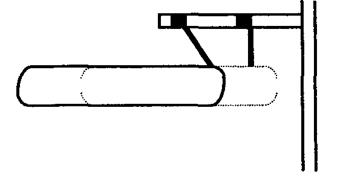
Малюнок 6.20-Скорочення м’язового волокна
Така кінематика рухів надмолекулярних структур при скороченні м'яза називається схемою ковзаючих ниток. Взаємодія міозинових| містків і ниток відбувається|походить| за рахунок енергії АТФ, що знаходиться|перебуває| у мітохондріях у присутності іонів кальцію.
Значення сили, що розвивається м’язами | при ізометричному скороченні, залежить від довжини саркомера, тобто від ступеню|міри| перекриття ниток (мал. 6.21):
o у м'язі, що знаходиться у спокої, містки не взаємодіють з|із| тонкою ниткою і його легко розтягнути. Розвинута сила при цьому дуже мала (до положення|становища| 1).
o від положення|становища| 2 кількість взаємодіючих містків збільшується, і сила, що розвивається м’язами|, зростає лінійно до максимального значення.
o від положення|становища| 2 до положення|становища| 5 сила взаємодії змінюється незначною мірою, оскільки кількість взаємодіючих містків практично не змінюється.
o від положення|становища| 5 до положення|становища| 6 сила взаємодії зменшується практично до нуля, оскільки подальше|дальше| зменшення саркомера приводить|призводить,наводить| до гальмування ковзання ниток і до деформацій товстих та тонких ниток.

Малюнок 621-Залежність сил скорочення м’язів від довжини саркомера
Таким чином, залежність сил скорочення м'язів від довжини саркомера можна розділити на 2 частини|частку|: перша частина|частка| від положення|становища| 1 до положення|становища| 2 — називається робочим циклом, і остання частина|частка| від положення|становища| 5 до положення|становища| 6 — холостим|холостим| циклом. Це подібно до двотактного двигуна внутрішнього згорання|згоряння|.
Слід зазначити також, що збудження групи м'язових волокон може здійснюватися від одного нервового волокна. У цьому випадку нервове волокно розгалужується на декілька синапсів. Сукупність м'язових волокон, які одночасно збуджуються (інервуються|) від одного нервового волокна, називається руховою одиницею. Різні рухові одиниці "розряджаються" (скорочуються) не одночасно, а хаотично, цим підтримується постійний тонус скелетного м'язу.
За своїми механічними властивостями м'язи відносяться до еластомерів — матеріалів, що володіють значною розтяжністю і пружністю.
Величина механічної напруги|напруження|, що виникає при скороченні м'язу (у певних межах, на малюнку 1.64. від положення|становища| 1 до положення|становища| 2), прямо пропорційна|пропорціональна| відносній деформації, згідно закону Гука.
 — механічна напруга|напруження|,
— механічна напруга|напруження|,  — сила, що розвивається,
— сила, що розвивається, 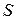 — площа|майдан| поперечного перетину;
— площа|майдан| поперечного перетину; 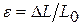 — відносна деформація,
— відносна деформація, 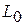 - початкова довжина,
- початкова довжина, 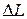 — абсолютна деформація (подовження|видовження|); Е — модуль пружності (модуль Юнга). Він визначається величиною| механічної напруги|напруження| у скороченому м'язі, при збільшенні її довжини в два рази.
— абсолютна деформація (подовження|видовження|); Е — модуль пружності (модуль Юнга). Він визначається величиною| механічної напруги|напруження| у скороченому м'язі, при збільшенні її довжини в два рази.
За своїми пружними властивостями та механізмом деформації м'язове волокно близько|поблизу| до каучуку: модуль пружності для обох матеріалів дорівнює 106Па. При сильній напрузі|напруженні| як у м'язі, так і у каучуку, спостерігається локальна кристалізація (впорядкування макромолекулярної білкової структури фіблярного типу).
Проте|однак|, при слабкому|слабому| і середньому скороченні, відновлення не буває повним|цілковитим|. Наявність залишкових деформацій характеризує пластичність м'язу — здатність|здібність| не зберігати свою форму і розміри після|потім| припинення дії сили. Таким чином м'яз не є|з'являється,являється| абсолютно пружним тілом, а має в'язкопружні властивості.
При дуже сильному розтягуванні м'яз поводиться як нормально пружне тіло. У цьому випадку температура м'язу знижується за рахунок локальної кристалізації.
При скороченні м'яза розвивається напруга|напруження| і здійснюється|скоюється,чиниться| робота. М'яз володіє скоротливими|скорочувальними| і еластичними елементами. Виникає напруга|напруження| і здійснюється робота, які обумовлені не тільки|не лише| активним скороченням скоротливого|скорочувального| комплексу, але і пасивним скороченням, що визначається еластичністю або так званим послідовним пружним компонентом м'яза. За рахунок послідовного пружного компоненту здійснюється|скоюється,чиниться| робота тільки|лише| у тому випадку, якщо|у тому випадку , якщо,в том случае | м'яз був заздалегідь розтягнутий, а величина цієї роботи пропорційна|пропорціональна| величині розтягування. Цим, більшою мірою, пояснюється те, що найбільш могутні рухи здійснюються|скоюються,чиняться| при великій амплітуді, що забезпечує попереднє розтягування м'язів.
М'язові скорочення діляться на ізометричні — м'язи, що відбуваються|походять| при незмінній довжині, і ізотонічні — що відбуваються|походять| при незмінній напрузі|напруженні|. В організмі скорочення м'язів ніколи не бувають чисто ізометричними або чисто ізотонічними.
Скелетні м'язи за допомогою сухожиль прикріпляються до кісток|кістей|, що утворюють систему важелів. Плече важеля м'яза, у більшості випадків, буває менше плеча важеля відповідної кістки|кісті|. Згідно Аккерману, механічний виграш в амплітуді рухів людини має величину від 2,5 до 20. Для двоголового м'яза плеча він дорівнює 10.
Залежно від сили, яку розвиває м'яз, швидкість скорочення (укорочення) м'яза буває різною. На основі достовірних|дослідних| даних, при роботі на ізольованих м'язах, Хілл вивів, так зване, основне рівняння скорочення м'яза. Згідно цього рівняння, швидкість скорочення м'яза знаходиться|перебуває| у гіперболічній залежності від величини навантаження F:
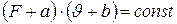 ,
,
де а і b — константи, рівні відповідно|відповідно до| 1/4F і ¼.
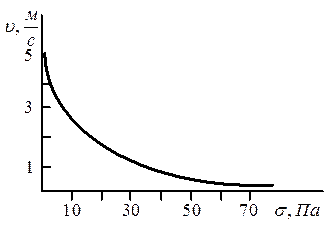
Малюнок 6.22 - Залежність швидкості скорочення м'яза від величини напруги|напруження|
Ця залежність швидкості скорочення м'яза від величини напруги|напруження| виражається|виказується,висловлюється| графічно (мал. 6.22).
Байером були зроблені зауваження до рівняння Хілла, що привели|призводять,наводять| рівняння до наступного|слідуючого| вигляду|виду|: 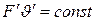 .
.
Добуток|добуток| 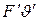 є загальною|спільною| потужністю, що розвивається м’язом при скороченні. Оскільки|тому що| F менша, тобто зовнішня потужність менша загальної|спільної| потужності, то слід припустити|передбачити|, що м'яз здійснює|скоює,чинить| не тільки|не лише| зовнішню роботу, але|та| збільшується на величину а, а швидкість скорочення — на величину b. Цю внутрішню роботу можна оцінювати як втрату енергії на внутрішнє тертя у формі теплового розсіяння. Тоді можна відзначити, що загальна|спільна| потужність м'язу, у фізіологічних межах, є|з'являється,являється| величиною постійного не залежного від величини навантаження і швидкості скорочення.
є загальною|спільною| потужністю, що розвивається м’язом при скороченні. Оскільки|тому що| F менша, тобто зовнішня потужність менша загальної|спільної| потужності, то слід припустити|передбачити|, що м'яз здійснює|скоює,чинить| не тільки|не лише| зовнішню роботу, але|та| збільшується на величину а, а швидкість скорочення — на величину b. Цю внутрішню роботу можна оцінювати як втрату енергії на внутрішнє тертя у формі теплового розсіяння. Тоді можна відзначити, що загальна|спільна| потужність м'язу, у фізіологічних межах, є|з'являється,являється| величиною постійного не залежного від величини навантаження і швидкості скорочення.
З термодинамічної точки зору, м'яз є системою, яка перетворює енергію хімічних зв'язків (енергію АТФ) в механічну роботу, тобто м'яз є|з'являється,являється| хемомеханічною| машиною. Як уже наголошувалося, при скороченні м'язу відбувається|походить| теплоутворення. Хіллом, за допомогою термоелектричних методів, було встановлено|установлено|, що при кожному роздратуванні на початку виділяється постійна по величині і незалежна від навантаження теплота активації Q, а потім теплота скорочення, пропорційна|пропорціональна| скороченню м'яза і незалежна від навантаження (до — коефіцієнт пропорційності).
Якщо скорочення ізотонічне, то м'яз виконує|виробляє,справляє| роботу А:
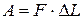
Згідно першого закону термодинаміки, зміна внутрішньої енергії 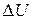 м'язу дорівнюватиме сумі виділеного тепла і звершеної|досконалої| роботи:
м'язу дорівнюватиме сумі виділеного тепла і звершеної|досконалої| роботи:
 або
або 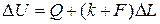
 або
або 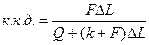
Враховуючи, що величини Q і до не залежать від F, з|із| рівняння виходить, що, в певних межах, ККД| м'язового скорочення збільшуватиметься при збільшенні навантаження.
Хілл, на підставі дослідних даних, визначив, що ККД| м'язового скорочення майже дорівнює 40%. Величина ККД| 40% показує ефективність перетворення енергії АТФ у механічну енергію.
Дата добавления: 2015-03-14; просмотров: 4923;