Електрогенез біопотенціалів
Між двома точками живої|жвавої| тканини за допомогою чутливої електровимірювальної апаратури можна зареєструвати постійну або змінну різницю потенціалів, які пов'язані з життєвою функцією організму і називаються біопотенціалами. Нижче наведені приклади|зразки| реєстрації біопотенціалів.
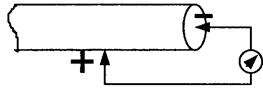
Малюнок 6.5-Потенціал пошкодження
1. Якщо розітнути живу|жваву| тканину, то між пошкодженою і непошкодженою|неушкодженою| ділянками можна зареєструвати постійну різницю потенціалів, яка називається демаркаційним потенціалом або потенціалом пошкодження|ушкодження| (мал. 6.5).
2. При накладенні електродів між внутрішнім і зовнішнім середовищами|середою| клітини|клітини| на поверхню нервових і м'язових волокон, які знаходяться|перебувають| в незбудженому стані, реєструється постійна різниця потенціалів, вона носить назву потенціал спокою (мал. 6.6).

Малюнок 6.6-Потенціал спокою Малюнок 6.7-Потенціал дії
1. Після нанесення роздратування на клітину|клітину|, нервове або м'язове волокно, вони переходять у збуджений стан. Зареєстрована різниця потенціалів, що при цьому змінюється, називається потенціалом дії(мал. 6.7).
2. Можна реєструвати різницю потенціалів окремих органів організму (мал. 6.8).
Для усіх наведених прикладів|зразків| спільним є|з'являється,являється| те, що вони пов'язані з життєвими процесами, які протікають у біологічних системах. Незважаючи на різні біопотенціали за своїми електричними характеристиками, всі вони так чи інакше обумовлені однією спільною причиною — нерівномірним розподілом іонів у двох різних ділянках тканини, тобто асиметрією концентрації іонів. Проте|однак| механізми їх утворення|утворення| у клітині можуть бути різні.
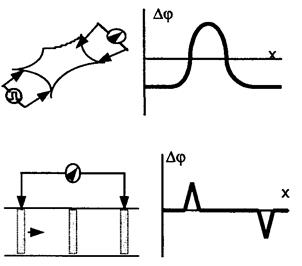
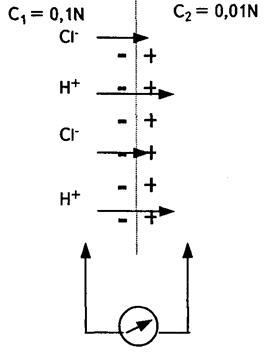
1. Дифузійний потенціал 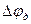 (мал. 6.9). Для його виникнення необхідний контакт електролітів з|із| різними концентрацією та рухливістю аніонів і катіонів
(мал. 6.9). Для його виникнення необхідний контакт електролітів з|із| різними концентрацією та рухливістю аніонів і катіонів 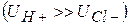 . На межі|кордон| розподілу для обох іонів характерне однакове проникнення.
. На межі|кордон| розподілу для обох іонів характерне однакове проникнення.
Перехід іонів 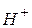 і
і 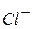 здійснюватиметься у бік меншої концентрації. Велика концентрація іонів водню з правого боку від межі|кордону| розділу електролітів буде створена внаслідок їх рухливості , тобто виникне різниця потенціалів . Кількісно величина визначається рівнянням Гендерсона:
здійснюватиметься у бік меншої концентрації. Велика концентрація іонів водню з правого боку від межі|кордону| розділу електролітів буде створена внаслідок їх рухливості , тобто виникне різниця потенціалів . Кількісно величина визначається рівнянням Гендерсона:
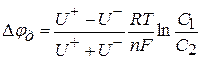
Малюнок 6.8-Реєстрація потенціалів окремих органів
2. Рівноважний мембранний потенціал 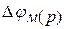 (мал. 6.10) Він виникає на межі|кордоні| розподілу двох електролітів, розділених мембраною, вибірково проникною для одного з іонів. Це випадок дифузійного потенціалу, коли рухливість одного з іонів
(мал. 6.10) Він виникає на межі|кордоні| розподілу двох електролітів, розділених мембраною, вибірково проникною для одного з іонів. Це випадок дифузійного потенціалу, коли рухливість одного з іонів
дорівнює нулю. При рухливості іонів рівній нулю, іони залишаються по один сторону бік мембрани, а іони проходять через мембрану, утворюючи на іншій стороні позитивний заряд.
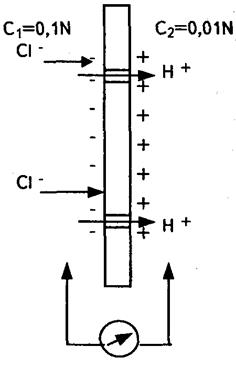
Малюнок 6..9- Дифузійний потенціал Малюнок 6.10-Рівноважний мембранний
потенціал
При цьому виникає потенціал, який описується рівнянням Ернста, окремий випадок рівняння Гендерсона, коли 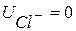 :
:
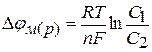
За рівнянням Нернста були розраховані мембранні потенціали клітин|клітини|, що створюються іонами Na+, K+, окремо|нарізно|. Проте|однак|, сумарний потенціал цих іонів значно відрізняється від експериментально визначеного|виміряного| на різних клітинах. Виникла необхідність створення|створіння| більш універсальної теорії, що пояснює створення біопотенціалів спокою клітини.
3. Стаціонарний мембранний потенціал 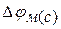 (мал. 6.11) Ходжкин і Катц припустили|передбачили|, що потенціал спокою клітин є|з'являється,являється| стаціонарним, тобто він обумовлений рухомою|жвавою,рухливою| рівновагою потоків іонів: Na+, K+, .
(мал. 6.11) Ходжкин і Катц припустили|передбачили|, що потенціал спокою клітин є|з'являється,являється| стаціонарним, тобто він обумовлений рухомою|жвавою,рухливою| рівновагою потоків іонів: Na+, K+, .
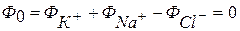
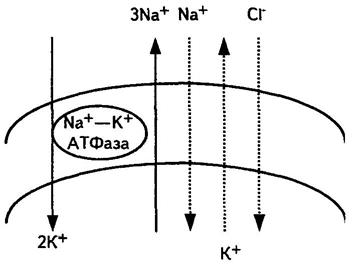
Малюнок 611- Стаціонарний мембранний потенціал
Сумарний потік, за їх припущенням,|гадках| обумовлений, з одного боку, активним транспортом іонів Na+ і К+ за рахунок енергії, що виділяється при гідролізі АТФ, з іншого боку, пасивним транспортом іонів Na+, K+ , оскільки|тому що| клітинна|кліткова| мембрана є проникненою для цих іонів. Вказані потоки постійні, згідно рівнянню Фіка, їх величини залежать від градієнта концентрації іонів на обох боках мембрани та від коефіцієнта проникності через пори і канали мембрани. Потенціал, що виникає за рахунок цього на мембрані, визначається рівнянням Гольдмана-Ходжкіна-Катца і називається стаціонарним:
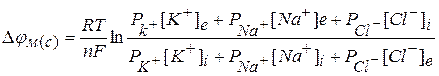
4. Потенціал електрогенної помпи  У попередньому завданні|задачі| було визначено стаціонарний потенціал на мембрані клітин|клітини|, що обумовлений сумарним потоком активного і пасивного транспорту іонів через мембрану. Проте|однак| можна розглядати|розглядувати| мембранний потенціал, виражений|виказаний,висловлений| тільки|лише| через характеристики активного транспорту речовин, зокрема через стехіометричний коефіцієнт
У попередньому завданні|задачі| було визначено стаціонарний потенціал на мембрані клітин|клітини|, що обумовлений сумарним потоком активного і пасивного транспорту іонів через мембрану. Проте|однак| можна розглядати|розглядувати| мембранний потенціал, виражений|виказаний,висловлений| тільки|лише| через характеристики активного транспорту речовин, зокрема через стехіометричний коефіцієнт  :
:
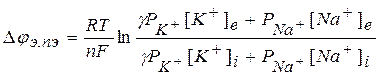 .
.
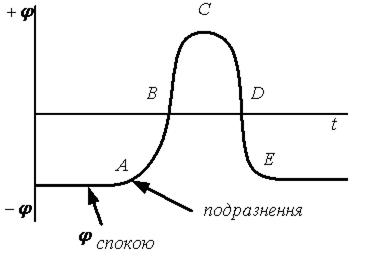
Малюнок 6.12-Механізм формування біопотенціалів дії
Таким чином, розглянуті|розгледіти| 4 випадки утворення постійної різниці потенціалів на мембранах. Усі ці випадки можуть приводити|призводити,наводити| до виникнення так званого потенціалу спокою на клітинній|клітковій| мембрані. Потенціал клітинної|кліткової| мембрани може змінюватися при дії різних природних або штучних подразників.
Розглянемо|розгледимо| механізм формування (генез) біопотенціалів дії (мал. 6.12)
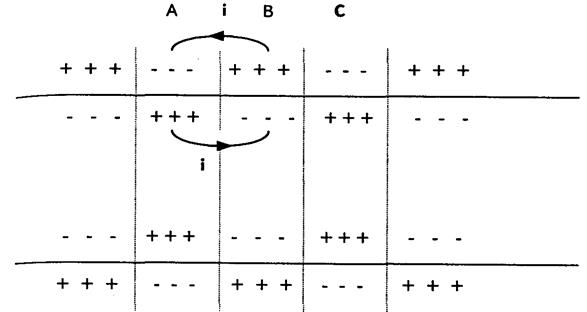
1. При роздратуванні клітин|клітини| (наприклад, прямокутними імпульсами) клітинна|кліткова| мембрана стає вибірково проникною для іонів Na+. Вони починають|розпочинають,зачинають| активно проникати всередину клітки|клітини| по градієнту, зменшуючи електронегативний потенціал протоплазми зрештою|врешті решт| 1 до 0 (лінія АВ|).
2. Подальше|дальше| надходження|вступ| Na+ у клітку|клітину| по градієнту інверсує| потенціал клітинної|кліткової| мембрани (лінія ВС|).
3. Досягши точки|точки| З|із| натрієві канали мембрани закриваються|зачиняються|, відкриваються|відчиняються| калієві канали. К+ починає|розпочинає,зачинає| активно виходити з|із| клітин|клітини| по градієнту, зменшуючи тим самим позитивний клітинний|клітковий| потенціал (лінія CD).
4. У точці|точці| D мембрана приходить в початковий|вихідний| стан по проникності для іонів К+ і Na+. Подальша|дальша| зміна потенціалу (лінія DE) відбувається|походить| за рахунок дії К+—Na+-помпи. У точці|точці| Е клітина приходить в початковий|вихідний| стан — збудження відсутнє.
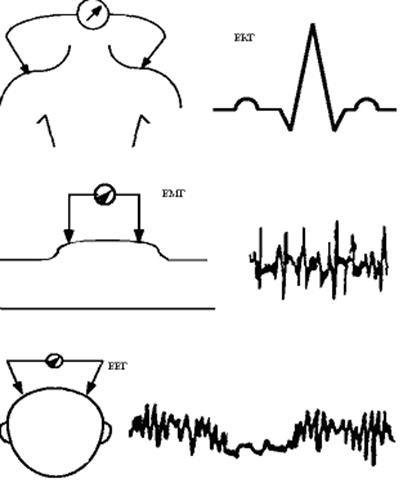
Для вимірювання|виміру| біопотенціалів спокою і біопотенціалів дії необхідна спеціальна апаратура (мал. 6.13).
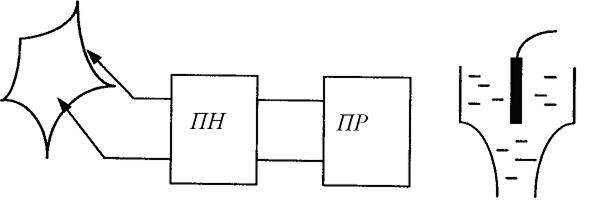
Малюнок 6.13-Апаратура для вимірювання біопотенціалів спокою і дії
1. Мікроелектроди. Вони виготовляються зі|із| скла і заповнюються децинормальним розчином КС1, у якому розміщається дріт з|із| хлорованого срібла. Діаметр мікроелектрода (0,1-1) мікрон, опір ~ 1 МОм, використовують тільки ті електроди, які не поляризуються.
2. Підсилювач напруги|напруження| біопотенціалів (ПН|) повинний мати дуже великий вхідний опір 10 МОм.
3. Як пристрої|устрої| реєстрації (ПР) можуть використовуватися осцилоскоп або самописець.
Наприкінці слід зазначити основні особливості формування біопотенціалів.
1. Первинною морфофункціональною| одиницею, в якій виникає джерело ЕРС|, є|з'являється,являється| клітина.
2. Біопотенціали, що створюються при роботі органів і тканин, є|з'являються,являються| результатом геометричної суми полів їх окремих клітин.
3. Біопотенціали мають іонну природу, їх причина — асиметрія концентрації іонів по обидва боки клітинної|кліткової| мембрани.
4. Біопотенціали відображають|відбивають| явища і процеси, що протікають у біологічних об'єктах, а електрограми є|з'являються,являються| одним з методів діагностики захворювань.
6.3 Активно-збудливі середовища|середа|
З|із| раніше розглянутого|розгледіти| відомо, що в незбудженому стані на мембрані живої клітини створюється постійна різниця потенціалів (потенціал спокою), яка обумовлена, в основному, рухомою|жвавою,рухливою| рівновагою іонів К+ Na+, , причому, зовнішня поверхня клітин|клітини| електропозитивна, внутрішня-електронегативна|. Кількісно ця модель описується рівнянням Гольдмана-Ходжкіна-Катца.
При збудженні, внаслідок зміни проникненості клітинних|кліткових| мембран для іонів Na+ і К+, відбувається|походить| спочатку деполяризація, а потім реполяризація збудженої ділянки клітинної|кліткової| мембрани, тобто виникає короткочасно змінний потенціал (потенціал дії).
Збудження триває декілька мілісекунд, потім мембрана повертається до початкового|вихідний| стану, але|та| різниця потенціалів на мембрані дорівнює нулю.
Відновлення потенціалу до початкового|вихідного| значення відбувається|походить| за рахунок калій-натрієвого| насосу.
Проте|однак|, збудження ділянки не є|з'являється,являється| локальним, воно переходить на інші ділянки, тобто процес збудження розповсюджується|поширюється| по поверхні клітин|клітини|. Математичний опис процесу розповсюдження|поширення| потенціалу дії було проведено по аналогії з фізичним процесом розповсюдження|поширення| електромагнітної хвилі по коаксіальному (ізольованому) кабелю.
Розглянемо|розгледимо| тільки|лише| принципові підходи до процесу розповсюдження|поширення| хвиль збудження у живих клітинах по аналогії з процесом розповсюдження|поширення| електромагнітних хвиль у середовищі|середі|.
Електромагнітна хвиля, розповсюджуючись|поширюючись| у середовищі|середі|, затухає,| оскільки її енергія переходить в енергію молекулярно-теплового руху середовища|середи|.
Хвилі збудження у живих|жвавих| тканинах не затухають, вони одержують|отримують| енергію із|із| самого середовища|середи| (енергію АТФ-фази), протягом періоду відновлення потенціалу спокою.
Хвилі, що одержують енергію із|із| самого середовища|середи| у процесі розповсюдження|поширення|, називаються автохвилями, а середовище|середа| називається активним, звідси і назва — активно-збудливі середовища|середа| (АЗС|).
Розглянемо|розгледимо| процес розповсюдження|поширення| збудження по нервовій клітині (нервовому волокну) мал. 6.14.
У збудженій ділянці А мембрана деполяризується і між збудженою А і незбудженою ділянками виникає різниця потенціалів 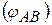 . Наявність різниці потенціалів приводить|призводить,наводить| до появи між цими ділянками електричних струмів, які називаються локальними струмами|токами| або струмами|токами| дії (i). На зовнішній ділянці поверхні клітин|клітини| локальний струм|тік| тече|теча| від не збудженої ділянки до збудженої, усередині клітин|клітини| — у зворотному напрямі. Локальний струм|тік| надає|робить,виявляє,чинить| дратівливу дію на незбуджену ділянку, викликаючи|спричиняючи| зменшення різниці потенціалів на цій ділянці, тобто деполяризацію мембрани. Коли деполяризація на ділянці В досягне порогового значення, у ньому виникає потенціал дії. В цей час на ділянці А відбуваються|походять| відновні процеси реполяризації, він стає не збудженим. Ділянка В, за описаним вище принципом, збуджує ділянку С і т.д.
. Наявність різниці потенціалів приводить|призводить,наводить| до появи між цими ділянками електричних струмів, які називаються локальними струмами|токами| або струмами|токами| дії (i). На зовнішній ділянці поверхні клітин|клітини| локальний струм|тік| тече|теча| від не збудженої ділянки до збудженої, усередині клітин|клітини| — у зворотному напрямі. Локальний струм|тік| надає|робить,виявляє,чинить| дратівливу дію на незбуджену ділянку, викликаючи|спричиняючи| зменшення різниці потенціалів на цій ділянці, тобто деполяризацію мембрани. Коли деполяризація на ділянці В досягне порогового значення, у ньому виникає потенціал дії. В цей час на ділянці А відбуваються|походять| відновні процеси реполяризації, він стає не збудженим. Ділянка В, за описаним вище принципом, збуджує ділянку С і т.д.
Це приводить|призводить,наводить| до розповсюдження|поширення| збудження по всій довжині нервового волокна, причому розповсюдження|поширення| відбувається|походить| тільки|лише| в одному напрямі|направленні|.
Насправді, в нервових волокнах відбувається|походить| односторонній|однобічний| напрям|направлення| розповсюдження|поширення| збудження за наявності мієлінових| оболонок, що покривають нервові волокна.
Малюнок 6.14-Процес поширення збудження по нервовому волокну
Оболонка є|з'являється,являється| ізолятором, і на ділянці, покритій мієліном, не може виникнути збудження. Мієлінова| оболонка не суцільна, через 1—3 мм вона порушується так званими перехопленнями Ранвьє — немієлізованими| ділянками (мал. 615).
При збудженні перехоплення А збуджується ділянка волокна тільки|лише| наступного|такого| перехоплення В, а якщо він блокований, наприклад анестетиком|, то наступним|слідуючим| збуджуватиметься перехоплення С. Після переходу збудження на наступне|таке| перехоплення В або С|із|, перехоплення А на деякий час втрачає|розгублює| здібність до збудження (властивість рефрактерності|) — цим обумовлене| одностороннє|однобічне| проведення розповсюдження|поширення| імпульсу по нервовому волокну.
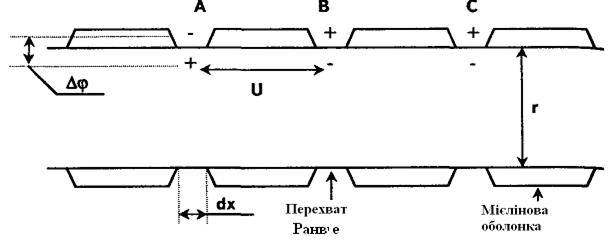
Малюнок 6.15- Перехвати Ранв'є
За електричним станом |збуджувані|ділянки нервового волокна, що зазнали збудження, можна представити|уявити| наступною|такою| еквівалентною схемою (мал. 6.16).
Тут: 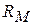 — опір ділянки мембрани на перехопленні Ранв'є.
— опір ділянки мембрани на перехопленні Ранв'є. 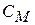 — ємність|ємкість| мембрани на перехопленні Ранв'є.
— ємність|ємкість| мембрани на перехопленні Ранв'є.
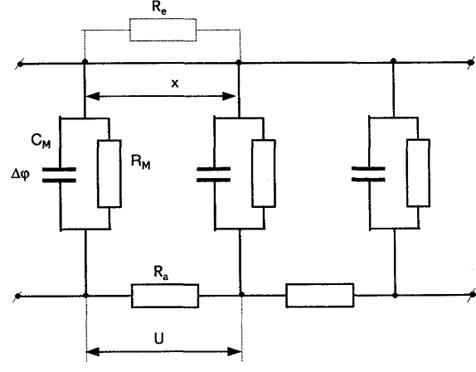
Малюнок 6.16-Електрична еквівалентна схема
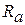 — опір аксоплазми| між перехопленнями Ранв'є. Опір зовнішнього середовища|середи|
— опір аксоплазми| між перехопленнями Ранв'є. Опір зовнішнього середовища|середи| 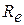 .
.
Позначимо величину потенціалу на мембрані при максимальній деполяризації 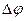 , заряд на мембрані у даний час
, заряд на мембрані у даний час 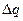 , ємність|ємкість| мембрани
, ємність|ємкість| мембрани 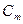 тоді
тоді 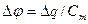 , звідки
, звідки 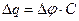 . Заряд мембрани та її потенціал змінюються унаслідок|внаслідок| протікання локального струму
. Заряд мембрани та її потенціал змінюються унаслідок|внаслідок| протікання локального струму 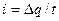 |току|, звідки
|току|, звідки 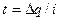 , де t — час, протягом якого мембрана деполяризується, тобто час переходу збудження з ділянки А на ділянку В або з ділянки В на ділянку С|із| і т.п. Локальний струм|тік| можна визначити з|із| формули
, де t — час, протягом якого мембрана деполяризується, тобто час переходу збудження з ділянки А на ділянку В або з ділянки В на ділянку С|із| і т.п. Локальний струм|тік| можна визначити з|із| формули 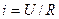 , де U — різниця потенціалів між збудженою і незбудженою ділянками, R — загальний|спільний| опір: мембрани
, де U — різниця потенціалів між збудженою і незбудженою ділянками, R — загальний|спільний| опір: мембрани 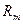 , аксоплазми
, аксоплазми 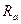 |, оточуючої клітку|клітину| рідини (останнім опором можна нехтувати зважаючи на|внаслідок,унаслідок| його невелике значення|крихта,малість|). Зрештою|врешті решт|
|, оточуючої клітку|клітину| рідини (останнім опором можна нехтувати зважаючи на|внаслідок,унаслідок| його невелике значення|крихта,малість|). Зрештою|врешті решт| 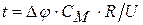 ;
; 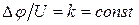 , тоді
, тоді 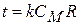 - це час переходу з однієї збудженої ділянки на іншу. Швидкість проведення збудження по нервовому волокну| дорівнює
- це час переходу з однієї збудженої ділянки на іншу. Швидкість проведення збудження по нервовому волокну| дорівнює 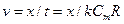 , де х — відстань між збудженою і не збудженою ділянками, а R дорівнює сумі опорів мембрани і аксоплазми:|
, де х — відстань між збудженою і не збудженою ділянками, а R дорівнює сумі опорів мембрани і аксоплазми:|
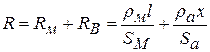 , де
, де
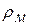 ,
, 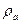 відповідно, питомі опори збудженої ділянки мембрани і аксоплазми|;
відповідно, питомі опори збудженої ділянки мембрани і аксоплазми|; 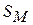 — товщина мембрани;
— товщина мембрани; 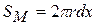 — площа|майдан| мембрани; r — радіус волокна, dx — ширина збудженої ділянки (перехоплення Ранв'є);
— площа|майдан| мембрани; r — радіус волокна, dx — ширина збудженої ділянки (перехоплення Ранв'є); 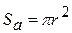 — площа|майдан| перетину аксоплазми|.
— площа|майдан| перетину аксоплазми|.
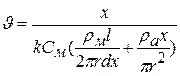 .
.
Остання формула визначає залежність швидкості розповсюдження|поширення| збудження по нервовому волокну від електричних параметрів — , 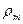 ,
, 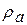 , геометричних розмірів волокна та збуджених ділянок — r, х,
, геометричних розмірів волокна та збуджених ділянок — r, х, 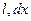 . Наприклад, якщо в мієліновому нервовому волокні блокувати один з каналів, то швидкість розповсюдження|поширення| збудження збільшується.
. Наприклад, якщо в мієліновому нервовому волокні блокувати один з каналів, то швидкість розповсюдження|поширення| збудження збільшується.
Ми розглянули|розгледіли| розповсюдження|поширення| збудження у активному провідному середовищі|середі| на прикладі|зразку| нервового волокна. Тепер звернемося|обернемося| до наймогутнішого джерела електричного поля в організмі людини — серцю.
Принципово будова|споруда| серця представлена|уявлена| на мал. 6.17. Серцевий|сердечний| пульс ритмічний і володіє певною частотою. Збудження у серці починається|розпочинається,зачинається| у сино-атріальному вузлі (венозному синусі) 1. Він діє подібно до релаксаційного генератора, що регулярно видає електричні імпульси (кожен через 1/78 хвилини). Ці імпульси розповсюджуються|поширюються| по поверхні передсердя на всіх напрямках, примушуючи|заставляючи| м'язові волокна передсердя скорочуватися.
Коли два імпульси приходять з протилежних напрямів і досягають протилежної сторони передсердя, |направлень|вони гасяться, оскільки|тому що| м'яз, що тільки що скоротився, не проводить імпульс у зворотному напрямі (рефрактерність|).
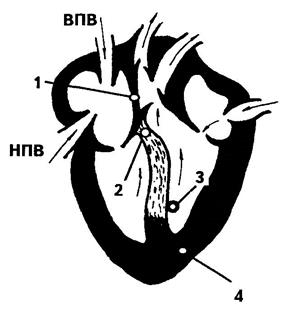
Малюнок 6.17-Серце
Крім того, імпульси примушують|заставляють| передсердя скорочуватися. Електрохімічна пульсація, зароджуючись у венозному синусі, стимулює, в першу чергу|передусім,насамперед| передсердно-шлуночковий (атріовентрикулярний) вузол 2. Цей вузол, після|потім| короткої затримки у часі (близько 0,1 сек) видає нові електричні імпульси, які розповсюджуються|поширюються| по спеціальній групі волокон, званих пучком Гіса (3). Ці волокна закінчуються у центральній м'язовій стінці між двома шлуночками. Імпульс розповсюджується|поширюється| далі по стінках шлуночка, досягаючи волокон Пуркіньє (4), та викликаючи|спричиняючи| скорочення шлуночків.
Сино-атріальний вузол відповідає електронному мультивібратору, що працює у режимі чекання і контролює інший мультивібратор — атріовентрикулярний вузол. Він, в свою чергу, контролює третій мультивібратор — шлуночок. Багато фактів свідчать на користь такої аналогії.
Власна частота сино-атріального| вузла може бути змодульована дією двох різних нервів — симпатикус| або парасимпатикус|, які знижують або підвищують частоту посилок|посилань| імпульсів на цей вузол. Це нагадує підстроювання опору або місткості|ємкості| у ланцюзі|цепі| працюючого мультивібратора.
Розглянувши фізико-хімічні|фізико-хімічні| принципи збудження у міокарді, слід зупинитися|зупинятися| на морфо-фізіологічних особливостях провідної системи серця. Вони полягають в тому, що кожна клітина |клітина|здатна|здібна| самостійно генерувати збудження, у цьому випадку говорять, що клітина|клітина| володіє автоматією. При цьому спостерігається градієнт автоматії, що виражається|виказується,висловлюється| в убуваючій здібності до автоматії у різних ділянок провідної системи, певною мірою віддалених|віддалення| від сино-атріального| вузла.
У звичайних|звичних| умовах автоматія усіх, розташованих нижче, ділянок пригнічується|придушується| частішими імпульсами із|із| сино-атріиального| вузла. У разі|в разі| поразки|ураження| або виходу з|із| ладу|строю,буд| цього вузла водієм ритму може стати антрио-вентрикулярний вузол. Імпульси при цьому виникатимуть з|із| частотою 30—40 за хвилину.
Якщо вийде з ладу|строю,буд| антрио-вентрикулярний вузол, то водієм ритму можуть стати волокна пучка Гіса. Частота знизиться до 15—20 за хвилину.
У тому випадку, коли|у тому випадку , якщо,в том случае | вийдуть з ладу|строю,буд| водії пучка Гіса, процес збудження спонтанно може виникнути у волокнах Пуркін'є. Ритм серця буде дуже низьким — до 10 за хвилину.
В цілому|загалом| діяльність серця можна представити|уявити| як роботу групи джерел струму|току| (струмових генераторів) в обмеженому провідному середовищі|середі|. Джерело струму|току| є ЕРС| з|із| внутрішнім опором r, що значно перевищує опір зовнішнього навантаження R.
Таким чином, струм|тік| у зовнішньому ланцюзі|цепі| залишається постійним незалежно від змін навантаження. Еквівалентна схема генератора струму|току| представлена|уявлена| на малюнку 6.18.
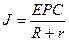 а оскільки|тому що|
а оскільки|тому що| 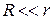
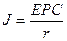
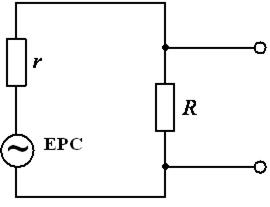
Малюнок 6.18-Еквівалентна схема
Залежно від частоти, форми та амплітуди напруги|напруження| джерела ЕРС| змінюється і струм|тік| у зовнішньому ланцюзі|цепі|, а отже і падіння напруги|напруження| на опорі R. Напруга|напруження| знімається із зручних точок зовнішнього ланцюга|цепу| і являє собою|з'являється,являється| ЕКГ, що відображає|відбиває| електричні та фізіологічні процеси, які відбуваються|походять| при розповсюдженні|поширенні| збудження у серці описаним вище способом. Тому ЕКГ і є|з'являється,являється| одним із засобів|коштів| діагностики захворювань серцево-судинної системи.
Дата добавления: 2015-03-14; просмотров: 2264;