Бактерии
Общая характеристика
Бактерии (прокариоты) — это большая группа микроорганизмов (около 1600 видов), большинство из которых одноклеточные.
Форма и размеры бактерий. Основные формы бактерий (рис. 2) — шаровидная, палочковидная и извитая.

Рис. 2. Форма бактерий:
а – шаровидные: 1 – микрококки, 2 – стрептококки, 3 — диплококки и тетракокки, – 4 стафилококки, 5 – сарцины;
б – палочковидные: 6 – палочки без спор, 7 – палочки со спорами;
в – извитые: 8 – вибрионы, 9 – спириллы, 10 – спирохеты;
г – редкие формы: 11 – тороиды, 12 – бактерии, образующие простеки, 13 – бактерии червеобразной формы, 14 – бактерии в форме шестиугольной звезды;
д – актиномицеты: 15 – мицелий, 16 – актиномицеты немицелиальные, 17 – споры
Шаровидные бактерии (рис. 2, а) — кокки имеют обычную форму шара, встречаются уплощенные, слабоовальной или бобовидной формы. Кокки могут быть в виде клеток одиночных — монококки (микрококки) или соединенных в различных сочетаниях: попарно — диплококки, по четыре клетки — тетракокки, в виде более или менее длинных цепочек — стрептококки, а также в виде скоплений кубической формы (в виде пакетов) из восьми клеток, расположенных в два яруса один над другим, — сарцины. Встречаются скопления неправильной формы, напоминающие грозди винограда, — стафилококки.
Палочковидные бактерии (рис. 2, б) могут быть одиночными или соединенными попарно — диплобактерии, цепочками по три-четыре и более клеток — стрептобактерии. Соотношения между длиной и толщиной палочек бывают самыми различными.
Извитые, илиизогнутые, бактерии (рис. 2, в) различаются длиной, толщиной и степенью изогнутости. Палочки, слегка изогнутые в виде запятой, называют вибрионами, палочки с одним или несколькими завитками в виде штопора — спириллами, а тонкие палочки с многочисленными завитками — спирохетами.
Благодаря использованию электронного микроскопа для изучения микроорганизмов в естественных природных субстратах были обнаружены бактерии, имеющие особую форму клеток (рис. 2, г): замкнутого или разомкнутого кольца (тороиды); с выростами (простеками); червеобразной формы — длинные с загнутыми очень тонкими концами; а также в виде шестиугольной звезды.
Размеры бактерий очень малы: от десятых долей микрометра (мкм) до нескольких микрометров. В среднем размер тела большинства бактерий 0,5—1 мкм, а средняя длина палочковидных бактерий — 2—5 мкм. Встречаются бактерии, размеры которых значительно превышают среднюю величину, а некоторые находятся на грани видимости в обычных оптических микроскопах.
Форма тела бактерий, как и их размеры, может изменяться в зависимости от возраста и условий роста. Однако при определенных, относительно стабильных условиях бактерии сохраняют присущие данному виду размеры и форму. Масса бактериальной клетки очень мала, приблизительно 4*10-14 г.
Строение бактериальной клетки. Клетка прокариотных организмов, к которым относятся бактерии, обладает принципиальными особенностями ультраструктуры. На рис. 3 представлена схема строения бактериальной клетки.
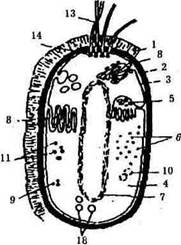
Рис. 3. Схема строения бактериальной клетки:
1 – капсула; 2 – клеточная стенка; 3 – цитоплазматическая мембрана; 4 – цитоплазма; 5 – мезосомы;
6 – рибосомы; 7 – нуклеоид; 8 – внутрицитоплазматические мембранные образования; 9 – жировые капли; 10 – полисахаридные гранулы; 11 – гранулы полифосфата; 12 – включения серы; 13 – жгутики; 14 – базальное тельце
Клеточная стенка (оболочка) — важный структурный элемент большинства бактерий (рис. 3, 2). На долю клеточной стенки приходится от 5 до 20% сухих веществ клетки. Она обладает эластичностью, служит механическим барьером между протопластом и окружающей средой, придает клетке определенную форму. В состав клеточной стенки входит специфическое для прокариотных клеток гетерополи-мерное соединение — пептидогликан (муреин), отсутствующий в клеточных стенках эукариотных организмов.
По методу окраски, предложенному датским физиком X. Грамом (1884 г.), бактерии делятся на две группы: грамположительные и грамотрицательные. Грамположительные клетки удерживают краску, а грамотрицательные не удерживают ее, что обусловлено различиями в химическом составе и ультраструктуре их клеточных стенок. У грамположительных бактерий клеточные стенки более толстые, аморфные, в них содержится большое количество муреина (от 50 до 90% сухой массы клеточной стенки) и тейхоевые кислоты. Клеточные стенки грамотрицательных бактерий более тонкие, слоистые, в них содержится много липидов, мало муреина (5-10%) и отсутствуют тейхоевые кислоты.
Клеточная стенка бактерий часто бывает покрыта слизью. Слизистый слой может быть тонким, едва различимым, но может быть и значительным, может образовыватькапсулу (рис. 3, 1). Нередко по размеру капсула намного превышает бактериальную клетку. Ослизнение клеточных стенок иногда бывает настолько сильным, что капсулы отдельных клеток сливаются в слизистые массы (зоогели), в которые вкраплены бактериальные клетки. Образуемые некоторыми бактериями слизистые вещества не удерживаются в виде компактной массы вокруг клеточной стенки, а диффундируют в окружающую среду. При быстром размножении в жидких субстратах слизеобразующие бактерии могут превратить их в сплошную слизистую массу. Такое явление наблюдается иногда в сахаристых экстрактах из свеклы при производстве сахара. За короткое время сахарный сироп может превратиться в тягучую слизистую массу. Ослизнению подвергаются мясо, колбасы, творог; наблюдается тягучесть молока, рассолов, квашеных овощей, пива, вина. Интенсивность слизеобразования и химический состав слизи зависят от вида бактерий и условий культивирования.
Капсула обладает полезными свойствами, слизь предохраняет клетки от неблагоприятных условий — у многих бактерий в таких условиях усиливается слизеобразование. Капсула защищает клетку от механических повреждений и высыхания, создает дополнительный осмотический барьер, служит препятствием для проникновения фагов, антител, иногда она является источником запасных питательных веществ.
Цитоплазматическая мембрана отделяет от клеточной стенки содержимое клетки (рис. 3,3). Это обязательная структура любой клетки. При нарушении целостности цитоплазматической мембраны клетка теряет жизнеспособность. На долю цитоплазматической мембраны приходится 8-15% сухого вещества клетки. В мембране содержится до 70-90% липидов клетки, толщина ее 7-10 нм. На срезах клеток в электронном микроскопе она видна в виде трехслойной структуры — одного липидного слоя и двух примыкающих к нему с обеих сторон белковых слоев. Цитоплазматическая мембрана местами впячивается внутрь клетки, образуя всевозможные мембранные структуры. В ней находятся различные ферменты; она полупроницаема, играет важную роль в обмене веществ между клеткой и окружающей средой.
Цитоплазма бактериальной клетки представляет собой полужидкую, вязкую, коллоидную систему (рис. 3, 4). Местами она пронизана мембранными структурами — мезосомами, которые произошли от цитоплазматической мембраны и сохранили с ней связь.
Мезосомы (рис. 3, 5) выполняют различные функции; в них и в связанной с ними цитоплазматической мембране имеются ферменты, участвующие в энергетических процессах — в снабжении клетки энергией. Хорошо развитые ме-зосомы обнаружены только у грамположительных бактерий, у грамотрицательных они развиты слабо и имеют более простое строение.
В цитоплазме содержатся рибосомы, ядерный аппарат и различные включения.
Рибосомы (рис. 3, 6) рассеяны в цитоплазме в виде гранул размером 20-30 нм; рибосомы состоят примерно на 60% из рибонуклеиновой кислоты (РНК) и на 40% из белка. Рибосомы ответственны за синтез белка клетки. В бактериальной клетке в зависимости от ее возраста и условий жизни может быть 5-50 тыс. рибосом.
Ядерный аппарат (рис. 3, 7). Электронная микроскопия ультратонких срезов через клетки бактерий позволила установить, что носителем генетической информации клетки является молекула дезоксирибонуклеиновой кислоты (ДНК). ДНК имеет форму двойной спиральной нити, замкнутой в кольцо; ее еще называют "бактериальная хромосома". Она расположена в определенном участке цитоплазмы, но не отделена от нее собственной мембраной. Этот ядерный аппарат бактериальных клеток называется нуклеоидом.
Цитоплазматические включения бактериальной клетки разнообразны, в основном это запасные питательные вещества, которые откладываются в клетках, когда они развиваются в условиях избытка питательных веществ в среде, и потребляются, когда клетки попадают в условия голодания. В клетках бактерий откладываются полисахариды: гликоген, крахмалоподобное вещество гранулеза, которые используются в качестве источника углерода и энергии. Липиды обнаруживаются в клетках в виде гранул и капелек. Жир служит хорошим источником углерода и энергии. У многих бактерий накапливаются полифосфаты; они содержатся в волютиновых гранулах и используются клетками как источник фосфора и энергии. В клетках серных бактерий откладывается молекулярная сера.
Подвижность бактерий. Шаровидные бактерии, как правило, неподвижны, Палочковидные бактерии бывают как подвижные, так и неподвижные. Изогнутые и спиралевидные бактерии подвижны. Движение большинства бактерий осуществляется с помощью жгутиков. Некоторые бактерии перемещаются путем скольжения.
Жгутики – это тонкие, спирально закрученные нити белковой природы, которые могут осуществлять вращательные движения. Длина жгутиков различна, а толщина так мала (10-20 нм), что в световой микроскопих можно увидеть только после специальной обработки клетки. Наличие, число и расположение жгутиков – постоянные для вида признаки и имеют диагностическое значение. Бактерии с одним жгутиком на конце клетки получили название люкотрихов, с пучком жгутиков – лофотрихов; с пучком жгутиков на обоих концах клетки – амфитрихов; бактерии, у которых жгутики находятся на всей поверхности клетки, называются перитрихами (рис. 4). Скорость передвижения велика: за секунду клетка со жгутиками может пройти расстояние в 20-50 раз больше, чем длина ее тела. При неблагоприятных условиях жизни, при старении клетки, при механическом воздействии подвижность может быть утрачена. Кроме жгутиков, на поверхности некоторых бактерий имеются в большом количестве нитевидные образования, значительно тоньше и короче, чем жгутики, – фимбрии (или пили).
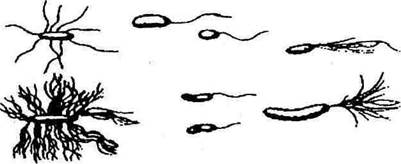
Рис. 4. Жгутики бактерий
Размножение бактерий. Для прокариотных клеток характерно простое деление клетки надвое. Деление клетки начинается, как правило, спустя некоторое время после деления нуклеоида. Палочковидные бактерии делятся поперек, шаровидные формы — в разных плоскостях. В зависимости от ориентации плоскости деления и их числа возникают различные формы: одиночные кокки, парные, цепочки, в виде пакетов, гроздьев.
Особенностью размножения бактерий является быстрота протекания процесса. Скорость деления зависит от вида бактерий, условий культивирования: некоторые виды делятся через каждые 15-20 мин, другие – через 5-10 ч. При таком делении число клеток бактерий за сутки достигает огромного количества. Это часто наблюдается на пищевых продуктах: быстрое скисание молока вследствие развития молочно-кислых бактерий, быстрая порча мяса и рыбы за счет развития гнилостных бактерий и т. д.
Спорообразование. Споры у бактерий образуются обычно при неблагоприятных условиях развития: при недостатке питательных веществ, изменении температуры, рН, при накоплении продуктов обмена выше определенного уровня. Способностью образовывать споры обладают в основном палочковидные бактерии. В каждой клетке образуется только одна спора (эндоспора).
Спорообразование – сложный процесс, в нем различают несколько стадий: сначала наблюдается перестройка генетического аппарата клетки, изменяются морфология нуклеоида. В клетке прекращается синтез ДНК. Ядерная ДНК вытягивается в виде нити, которая затем разделяется; часть ее концентрируется у одного из полюсов клетки. Эта часть клетки называется спорогенной зоной. В спорогенной зоне происходит уплотнение цитоплазмы, затем этот участок обособляется от остального клеточного содержимого перегородкой (септой) (рис. 5, а; I-IV). Отсеченный участок покрывается мембраной материнской клетки, образуется так называемая проспора. Простора — это структура, располагающаяся внутри материнской клетки, от которой она отделена двумя мембранами: наружной и внутренней (рис. 5, а; III-IV). Между мембранами формируется кортикальный слой (кортекс), сходный по химическому составу с клеточной стенкой вегетативной клетки. Помимо пептидогликана, в кортексе содержится дипиколиновая кислота (С7Н8О4N), которая отсутствует в вегетативных клетках. В дальнейшем поверх просторы образуется оболочка споры, состоящая из нескольких слоев. Число, толщина и строение слоев различны у разных видов бактерий. Поверхность наружной оболочки может быть гладкой либо с выростами разной длины и формы. Поверх оболочки споры нередко образуется еще тонкий покров, окружающий спору в виде чехла, – экзоспориум (рис. 5, б; 1).
Споры имеют обычно круглую или овальную форму. Диаметр спор некоторых бактерий превышает ширину клетки, вследствие чего форма спороносящих клеток, изменяется. Клетка приобретает форму веретена (клостридиум), если спора расположена в ее центре, или форму барабанной палочки (плектридиум), когда спора приближена к концу клетки.
После созревания споры материнская клетка отмирает, оболочка ее разрушается и спора освобождается. Процесс образования споры протекает в течение нескольких часов.
Наличие у бактериальных спор плотной, труднопроницаемой оболочки, малое содержание в ней воды, большое количество липидов, а также наличие кальция и дипиколиновой кислоты обусловливают высокую устойчивость спор к факторам внешней среды. Споры могут находиться в жизнеспособном состоянии сотни и даже тысячи лет. Например, жизнеспособные споры выделены из трупов мамонтов и египетских мумий, возраст которых исчисляется тысячелетиями. Споры устойчивы к высокой температуре: в сухом состоянии они погибают после прогревания при 165-170°С в течение 1,5-2 ч, а при перегретом паре (в автоклаве) – при 121°С в течение 15-30 мин.
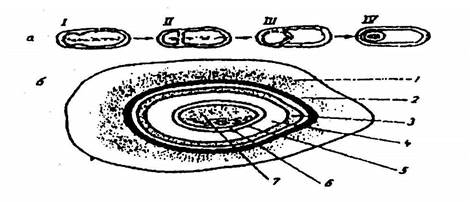
Рис. 5. Цикл развития (а) и схема строения зрелой бактериальной споры (б):
I-II – процесс отделения спорогенной зоны; III-IV – образование проспоры; 1 – экзоспориум; 2 – наружная оболочка споры; 3 – внутренняя оболочка споры; 4 – кортекс; 5 – клеточная стенка зародыша; 6 – цитоплазматическая мембрана; 7 – цитоплазма с ядерным веществом
В благоприятных условиях спора прорастает в вегетативную клетку; этот процесс обычно длится несколько часов.
Прорастающая спора начинает активно поглощать воду, активизируются ее ферменты, усиливаются биохимические процессы, приводящие к росту. Кортекс при прорастании споры превращается в клеточную стенку молодой вегетативной клетки; освобождаются во внешнюю среду дипиколиновая кислота и кальций. Внешняя оболочка споры разрывается, через разрывы выходит наружу "росток" новой клетки, из которого затем формируется вегетативная бактериальная клетка.
Порчу пищевых продуктов вызывают лишь вегетативные клетки. Знание факторов, способствующих образованию спор у бактерий, и факторов, которые вызывают их прорастание в вегетативные клетки, имеет значение в выборе способа обработки продуктов с целью предотвращения их микробной порчи.
Изложенные выше сведения характеризуют в основном так называемые истинные бактерии. Существуют и другие, более или менее отличающиеся от них, к которым относятся следующие.
Нитчатые (нитевидные бактерии). Это многоклеточные организмы в виде нитей различной длины, диаметром от 1 до 7 мкм, подвижных или прикрепленных к субстрату. В основном нити со слизистым чехлом. Они могут содержать окись магния или окислы железа. Живут в водоемах, встречаются в почве.
Миксобактерии. Это палочковидные бактерии, передвигаются путем скольжения. Они образуют плодовые тела – скопления клеток, заключенных в слизь. Клетки в плодовых телах переходят в покоящееся состояние – миксо-споры. Эти бактерии живут в почве, на различных растительных остатках.
Почкующиеся и стебельковые бактерии размножаются почкованием, образуют стебельки или то и другое вместе. Есть виды с выростами - простеками (см. рис. 1, 12).Живут в почве и водоемах.
Актинолшиеты. Бактерии имеют ветвистую форму. Одни — палочки слегка разветвленные (см. рис. 2, д), другие – в виде тонких ветвящихся нитей, образующих одноклеточный мицелий. Мицелиальные актиномицеты, называемые "лучистые грибки", размножаются спорами, развивающимися на воздушных ветвях мицелия. Актиномицеты бывают окрашены; они широко распространены в природе. Встречаются и на пищевых продуктах и могут вызвать их порчу. Продукт приобретает характерный землистый запах. Многие актиномицеты продуцируют антибиотики. Есть виды, патогенные для человека и животных.
Риккетсии. Палочковидные и кокковидные микроорганизмы, неподвижны, спор не образуют. Внутриклеточные паразиты; некоторые вызывают заболевания животных, и человека. Переносчиками риккетсий в основном являются насекомые (вши, блохи, клещи).
Микоплазмы. Организмы без клеточной стенки, покрыты лишь трехслойной мембраной.
Клетки очень мелкие, иногда ультрамикроскопических размеров (около 200 нм), плеоморфные (разнообразной формы) – от кокковидных до нитевидных. Некоторые вызывают заболевания человека, животных, растений.
Основы систематики бактерий
Систематика (таксономия) организмов заключается в распределении (классификации) их по определенным группам, каждая из которых имеет название: класс, порядок, семейство, род, вид. Вид – основная таксономическая единица.
В микробиологии часто употребляют термин "штамм". Это более узкое понятие, чем вид. Штаммами называют чистые культуры микроорганизмов одного и того же вида, выделенные из различных сред (субстратов).
В соответствии с принятыми в биологии правилами название бактерий, как и других организмов, дается на латинском языке и состоит из двух слов, первое обозначает
род, к которому принадлежит данная бактерия, второе – вид. Родовое название пишется с прописной буквы, видовое – со строчной. Например, 81;гер1;ососсиз 1ас11з относится к шаровидным бактериям, образующим цепочки (род 51;гер1.ососси5), они вызывают скисание молока в результате сбраживания сахара в молочную кислоту, отсюда и видовое название 1ас^з.
С 1 января 1980 г. введен новый кодекс номенклатуры бактерий[1]. Названия бактерий в учебнике даются в соответствии с новой номенклатурой[2].
Систематика бактерий – сложная проблема. Единой "естественной" (филогенетической) классификации их, отражающей родственные связи между отдельными группами бактерий, эволюционное развитие отдельных видов, не существует.
Современные системы классификации бактерий по существу являются искусственными, объединяют бактерии в определенные группы на основе сходства их по комплексу морфологических, физиологических, биохимических признаков (в частности, по составу ДНК). Эти системы используются для распознавания (определения – идентификации) тех или иных бактерий.
В этих целях наиболее широко используется "Руководство Берги по определению бактерий" (1974, 8-е издание). У нас оно было издано в 1980 г. в сокращенном виде "Краткий определитель бактерий Берги".
В указанных "Руководствах" все прокариотические микроорганизмы объединены в царство прокариот (Ргокагуо1ае), которое подразделено на два отдела – цианобактерии и бактерии.
I отдел. Цианобактерии (сине-зеленые водоросли). Это фототрофные микроорганизмы. Некоторые цианобактерии одноклеточны, другие – многоклеточны, образуют простые или ветвящиеся нити. Клетки многих покрыты слизистой
капсулой. Обитают цианобактерии преимущественно в водоемах. Эти бактерии привлекают внимание как продуценты пищевого белка.
II отдел. Бактерии. Этот отдел разделен на 19 частей (групп), каждая из которых делится на порядки, порядки – на семейства, семейства – на роды, роды – на виды.
В 1984 г. вышло новое, 9-е издание "Руководства Берги по систематике бактерий". В нем царство прокариот (Ргокагуо1ае) подразделено на четыре отдела в зависимости от наличия или отсутствия клеточной стенки и ее состава.
В первый отдел – тонкокожих – включены группы бактерий, относящихся к окраске по Граму отрицательно, а также фототрофные бактерии и цианобактерии.
Во второй отдел – твердокожих – включены группы бактерий, относящихся к окраске по Граму положительно.
В третий отдел включены микоплазмы – бактерии, не имеющие клеточной стенки.
В четвертый включены метанообразующие бактерии и особая группа бактерий, имеющих несовершенную клеточную стенку: в ней нет характерного для прокариот муреина, а содержатся другие биополимеры. Как показали исследования последних лет, эти бактерии значительно отличаются от всех остальных по морфологическим, физиологическим, биохимическим и цитологическим признакам. Эту особую группу бактерий назвали архебактериями (Аrсhеоbасtеriа).
Форма их клеток разнообразна: шар, цилиндр, луч, звезда, квадрат, коробочка и др. Как и у других прокариот, генетическим аппаратом является нуклеоид, но в цитоплазме имеются характерные для эукариот мембранные органеллы (митохондрии, эндоплазматический ретикулум и др.). Большинство из них строгие анаэробы, но есть и аэробы, гетеротрофы и автотрофы. Они обитают в экстремальных условиях внешней среды. Многие из них крайне термофильны – температурный оптиум 85-105°С, обнаружены в горячих источниках. Некоторые крайне галофильны: выделены из солеварен, где добывается морская соль. Есть и крайне ацидофильные, способные развиваться при рН 1-2. По-видимому, архебактерии – одна из древнейших форм жизни.
Ознакомление с указанными "Определителями" при идентификации бактерий проводится на лабораторных занятиях.
Вирусы и фаги
Вирусы (от лат. virus – яд) – это особая группа микроорганизмов меньших размеров и более простой организации, чем бактерии. Вирусы не имеют клеточной структуры, величина их измеряется нанометрами. Вирусы открыты русским ботаником Д. И. Ивановским в 1892 г. при изучении мозаичной болезни листьев табака, которая причиняла большой ущерб табачным плантациям Крыма. Открытие Д. И. Ивановского заложило основу новой науки – вирусологии.
Вирусы – внутриклеточные паразиты, вызывающие многие болезни человека (оспу, грипп, бешенство, корь, полиомиелит и др.), животных (ящур, чуму крупного рогатого скота) и растений ("мозаики" и другого вида заболевания полевых и огородных культур).
Вирусы разнообразны по форме, размерам и химическому составу. Большинство из них имеет палочковидную или сферическую форму (рис. 6, а). Некоторые вирусы состоят только из белка и одной нуклеиновой кислоты – ДНК или РНК, другие содержат еще и липиды, полисахариды. Вирусная частица называется вирионож Нуклеиновая кислота (в виде спирали) находится внутри вириона, снаружи он покрыт белковой оболочкой (капсидом), состоящей из отдельных морфологических субъединиц (капсомеров) (рис. 6, б). Вирусы выращивают на живых клетках или культуре тканей, так как на искусственных питательных средах они, как правило, не развиваются.
Вирусы обладают разной устойчивостью к внешним воздействиям. Многие инактивируются при 60"С до 10 мин, другие выдерживают температуру 90°С до 10 мин. Вирусы довольно легко переносят высушивание и низкие температуры, но мало устойчивы ко многим антисептикам, ультрафиолетовым лучам, радиоактивным излучениям.
Фаги– это вирусы микроорганизмов, вызывающие гибель – распад (лизис) их клеток. Вирусы бактерий называются бактериофагами или просто фагами, актиномицетов – актинофагами, вирусы грибов – микофагами, сине-зеленых водорослей (цианобактерий) – цианофагами.
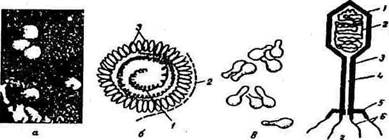
Рис. 6. Электронная микроскопия вируса и бактериофага;
а – вирус оспенной вакцины;
б – схематическое изображение простого вириона: 1 – нуклеиновая кислота, 2 – капсид, 3 – капсомеры;
в – бактериофаг; г – схема строения фага: 1 – головка, 2 – ДНК, 3 – отросток, 4 – стержень, 5 – пластинка отростка, 6 – нити
Впервые лизис сибиреязвенных бактерий наблюдал Н. Ф. Гамалея в 1898 г. Д'Эррель в 1917 г. установил явление лизиса у бактерий дизентерии, им впервые был выделен и описан бактериофаг ("пожиратель") бактерий.
Морфология фага (рис. 6, в) изучена с применением электронного микроскопа. Большинство фагов состоит из головки и отростка. Головка фага может иметь разную форму, чаще всего это многогранник, покрытый белковой оболочкой (капсидом). Внутри капсида расположена нуклеиновая кислота, чаще всего одна – ДНК или РНК. Отросток фага имеет внутренний полый стержень, по каналу которого ДНК фага переходит в клетку хозяина. Стержень снаружи покрыт чехлом, способным к сокращению. Стержень и чехол отростка состоят из белковых субъединиц. У некоторых фагов отросток заканчивается базальной пластинкой, которая имеет выступы (зубцы) и нити.
Фаги могут быть и нитевидной формы, могут состоять из одной головки, а могут быть с аналогами отростка (очень коротким отростком). Некоторые фаги имеют длинные отростки с несокращающимся или сокращающимся чехлом.
Фаги широко распространены в природе. Многие из них обладают специфичностью – могут воздействовать на определенный вид или группу родственных видов микроорганизмов.
Взаимодействие фага с микробной клеткой происходит в несколько фаз. Сначала фаг адсорбируется восприимчивой клеткой, затем под действием фермента фага (сходного с лизоцимом) в стенке микробной клетки образуется отверстие, через которое в клетку проникает только нуклеиновая кислота; пустая белковая оболочка головки и отростка остается снаружи клетки, а затем разрушается.
Под влиянием попавшей в клетку нуклеиновой кислоты фага перестраиваются все обменные процессы микробной клетки на синтез фаговых частиц: синтезируются фаговая нуклеиновая кислота и белковые субъединицы оболочек. Вначале формируются раздельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. Через определенное время клетка хозяина погибает, разрушается, и фаги выходят наружу.
Явление фаголизиса (растворение культур микроорганизмов) наблюдается на производствах, связанных с использованием микроорганизмов. Развитие фагов в культурах промышленных микроорганизмов приводит к тому, что клетки культуры лизируются, не успев синтезировать необходимые вещества. Это наносит предприятиям большой экономический ущерб. Так нередко лизируются молочно-кислые бактерии, входящие в состав заквасок для кисло-молочных продуктов. Такие закваски не пригодны для употребления.
Бактериофаги, лизирующие зараженные ими бактерии, называют вирулентными. Некоторые фаги, Однако, инфицируют бактерии, но не вызываютих лизиса; такие фаги называются умеренными. В клетке-бактерии хозяина они не размножаются, но при делении бактерии передаются дочерним клеткам.
Фаги применяются в медицине для лечения и профилактики некоторых заболеваний, например дизентерии, холеры. Фаги исключительно удобны как модели для решения вопросов общебиологических, молекулярной биологии, генетики, медицины.
Грибы
Общая характеристика
Грибы (Мусоtа) — обширная и разнообразная группа растительных организмов. Они не содержат хлорофилла и не способны к синтезу органических веществ из углекислого газа; грибы – хемоорганотрофы. В природе грибы обитают на разнообразных субстратах, в почве, в воде и играют важную роль в круговороте веществ в природе.
Многие грибы употребляют в пищу, используют в промышленных условиях для получения органических кислот, витаминов, ферментов, антибиотиков.
Многочисленные грибы, развивающиеся на пищевых продуктах, промышленных материалах и изделиях, вызывают их порчу и разрушение. Некоторые из них способны вырабатывать токсические для человека и животных вещества – микотоксины. Многие грибы поражают культурные растения в процессе их вегетации, нанося большой урон сельскому хозяйству. Есть грибы, вызывающие заболевания человека и животных.
Строение тела гриба. Вегетативное тело большинства грибов представляет собой грибницу, или мицелий, состоящий из ветвящихся нитей – гиф. Такие грибы называют мицелиальными (еще их называют плесенями).
С помощью сканирующего электронного микроскопа установлено (А. А. Кудряшова), что гифы грибов различаются внешним видом, строением стенки, длиной, толщиной и рельефом поверхности. Они могут быть прямыми, изогнутыми, спиралевидными, со вздутиями или утолщениями, с углублениями и короткими отростками "корешками", служащими для прикрепления к субстрату. Поверхность гиф бывает с шипами, гладкой, сетчатой, волокнистой, местами складчатой (рис. 7). Диаметр гиф колеблется от 2 до 25 мкм и более.

Рис. 7. Электронная микрофотография рельефа поверхности гиф
1 – Rizopus nigricans; 2 – Оidium lactus; 3 — Реnicillum ехраnsum
Гифы растут вершиной или концами разветвлений, поэтому их клетки неоднородны по длине. Мицелий развивается частично в субстрате (субстратный мицелий), пронизывая его и высасывая из него воду и питательные вещества, а частично — на поверхности субстрата (воздушный мицелий) в виде пушистых, паутинообразных или тонких налетов, пленок. Гифы отдельных грибов могут плотно переплетаться и даже срастаться между собой. У некоторых грибов гифы соединяются параллельно в тяжи, достигающие иногда нескольких метров в длину, по ним притекают питательные вещества.
Немногие грибы не имеют мицелия. Это некоторые представители низших грибов, а также дрожжи, которые являют собой одиночные округлые или удлиненные клетки.
Мицелий одних грибов клеточный – гифы разделены перегородками (септами) на клетки, часто многоядерные; мицелий других – неклеточный, гифы не имеют перегородок, и весь мицелий представляет собой как бы одну гигантскую клетку с большим числом ядер (рис. 8).
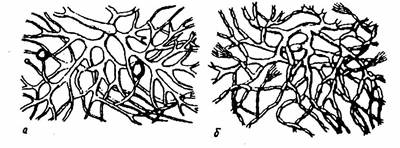
Рис. 8. Мицелий грибов:
а – неклеточный; б – клеточный
Из плотного сплетения гиф состоят так называемые плодовые тела грибов, в которых находятся органы размножения.
Видоизмененным мицелием являются склероции – обычно темные, различной формы образования из плотно переплетенных гиф (см. рис. 10, б).
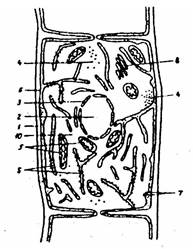
Рис. 9. Схема строения грибной клетки:
1 – клеточная стенка; 2 – ядро; 3 – ядерная оболочка; 4 – рибосомы; 5 – митохондрии; 6 – цитоплазматическая мембрана; 7 – лизосомы; 8 – аппарат Гольджи; 9 – эндоплазматическая сеть; 10 – цитоплазма; 11 – вакуоль
Склероции устойчивы к неблагоприятным условиям внешней среды, богаты запасными питательными веществами.
Строение клетки. Грибы имеют эукариотный тип клетки, строение которой сходно с клетками других растительных организмов, но у грибов отсутствуют пластиды (рис. 9).
Клетки большинства грибов имеют многослойную клеточную стенку, состоящую на 80-90% из полисахаридов; в небольшом количестве имеются белки, липиды, полифосфаты.
Основным полисахаридом клеточной стенки большинства грибов является хитин, у некоторых – целлюлоза. Под клеточной стенкой расположена трехслойная цитоплазматическая мембрана.
В цитоплазме находятся многочисленные органоиды – структуры различного строения и функций.
Митохондрии – образования из липопротеиновых мембран, в которых осуществляются энергетические процессы и синтезируется АТФ-вещество, богатое энергией.
Эндоплазматический ретикулум (эндоплазматическая сеть) – мембранная система из взаимосвязанных канальцев (местами суживающихся или расширяющихся), которая пронизывает цитоплазму и связана с цитоплазматической мембраной и мембраной ядра. В этом органоиде происходит синтез многих веществ (липидов, углеводов и др.).
Аппарат Гольджи – мембранная система, связанная с ядерной мембраной и с эндоплазматической сетью. К его многообразным функциям относятся транспортирование веществ, синтезируемых в эндоплазматической сети, а также удаление из клетки продуктов обмена.
Рибосомы – очень мелкие, округлые, многочисленные образования. Часть их находится в свободном состоянии, часть прикреплена к мембранам. В рибосомах происходит синтез белка.
Лизосомы – мелкие округлые тельца, покрытые мембраной. В них содержатся ферменты, переваривающие (расщепляющие) поступающие извне белки, углеводы, липиды.
Ядро (или несколько ядер) окружено двойной мембраной. В нуклеоплазме имеются ядрышко и хромосомы, содержащие ДНК. В ядерной оболочке расположены поры, обеспечивающие транспорт веществ между ядром и цитоплазмой.
Вакуоли – полости, окруженные мембраной, заполненные клеточным соком и включениями запасных питательных веществ (волютина, гликогена, жира).
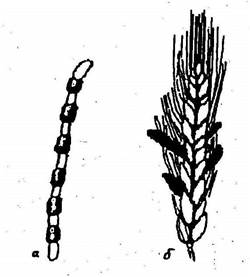
Рис. 10. Хламидоспоры и склероции грибов:
а — хламидоспоры; б — склероции спорыньи
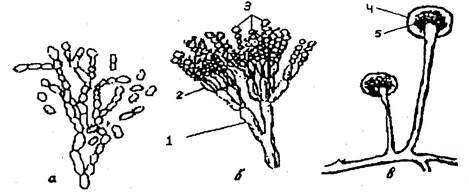
Рис. 11. Органы бесполого размножения грибов:
а – оидии; б – конидиеносец (1) со стеригмами (2) и конидиями (3); в – спорангиеносец со спорангием (4) и спорангиоспорамй (5)
Размножение грибов. Особенностью грибов является большое разнообразие способов и органов размножения. Один и тот же гриб часто имеет несколько форм размножения. При этом внешний вид гриба может настолько изменяться, что каждом из них гриб рассматривают как самостоятельный вид.
Грибы размножаются вегетативным, бесполым и половым путями.
Вегетативное размножение происходит без образования каких-либо специализированных органов: частями мицелия или отдельными клетками оидиями (артроспорами), образующимися в результате расчленения гиф (рис. 11, а), которые на питательном субстрате разрастаются в грибницу. Размножение происходит и образующимися на гифах хламидоспорами (рис. 10, а) – толстостенными клетками, устойчивыми к неблагоприятным условиям.
При бесполом и половом размножении образуются специализированные клетки – споры, с помощью которых и осуществляется размножение.
Прибесполом способе размножения споры образуются на особых гифах воздушного мицелия, внешне отличающихся от других гиф.
У одних грибов споры образуются экзогенно (открыто) – на вершине гиф снаружи их. Такие споры называются конидиями, а гифы, несущие их, – конидиеносцами (рис. 11, б).
Конидиеносцы развиваются на мицелии поодиночке или группами. При групповом развитии конидиеносцы одних грибов объединяются в пучки (коремии), у других они располагаются тесным слоем в особых кувшиновидных (пикниды) или блюдцеобразных (ложе) образованиях из плотного сплетения гиф (рис. 12).
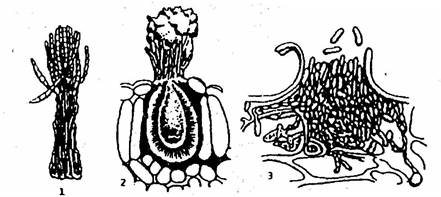
Рис. 12. Типы конидиального спороношения:
1 – коремия; 2 – пикнида; 3 – ложе
Конидии образуются непосредственно на конидиеносце или на специальных клетках, расположенных на его вершине. Эти клетки обычно имеют форму бутылочек и называются стеригмами или фиалидами. Конидии располагаются на конидиеносцах (или на стеригмах) поодиночке, группами, цепочками и т. д.
У других грибов споры образуются эндогенно – внутри особых клеток, развивающихся на концах гиф. Эти клетки – вместилища спор – называются спорангиями, находящиеся в них споры – спорангиоспорамй, а гифы, несущие спорангии со спорами, – спорангиеносцами (рис. 11, в). От несущей гифы спорангий отделен перегородкой (колонкой), врастающей внутрь спорангия.
У некоторых грибов в спорангиях образуются подвижные споры, снабженные жгутиками, — зооспоры.
Спорангиоспоры и конидии бывают различной формы, размера и окраски, благодаря чему грибы в стадии спороношения имеют вид окрашенных налетов. Созревшие конидии осыпаются. При созревании спорангиоспор спорангии лопаются и из них высыпаются споры. Конидии и спорангиоспо-ры пассивно разносятся потоками воздуха на большие расстояния. Попав в благоприятные условия, споры прорастают в гифы.
Спорангиеносцы, и особенно конидиеносцы грибов, имеют разнообразное строение и внешний вид, типичные для отдельных представителей.
Приполовом размножении грибов спорообразованию предшествует половой процесс — слияние половых клеток с последующим объединением их ядер. В результате образуются специализированные органы размножения. Развитие этих органов, формы полового процесса у грибов многообразны.
У грибов с клеточным мицелием в качестве органа полового размножения образуются базидии со спорами или сумками со спорами.
Базидия представляет собой мешковидно вытянутую клетку, на которой имеются выросты-стеригмы (обычно четыре), на каждом из которых находится по одной споре. Эти споры называются базидиоспорами (рис. 13, а, 1). Базидии бывают и многоклеточными (рис. 13, а, 2).
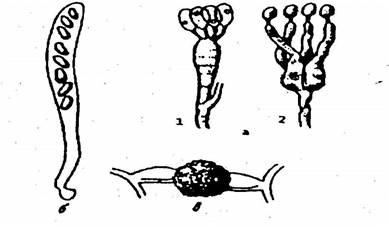
Рис. 13. Органы полового спороношения:
а – базидии с базидиоспорами: 1 – одноклеточная базидия, 2 – многоклеточная базидия; б – сумка (аскус) с аскоспорами; в – зигоспора
Сумка (аскус) имеет вид цилиндрической клетки, внутри которой находятся споры (чаще восемь), называемые аскоспорохи (рис. 13, б). Аскоспоры бывают различной формы, бесцветными или окрашенными.
Базидии и сумки иногда располагаются на мицелии поодиночке, но большей частью они развиваются группами или слоями в особых образованиях из плотно переплетенных гиф – плодовых тел. По форме, строению и окраске плодовые тела очень разнообразны. Такими плодовыми телами являются, например, шляпка с ножкой белого гриба, сыроежки, опенка и др.
У грибов с неклеточным мицелием в результате полового процесса образуется одна спора – зигоспора или ооспора (рис. 13, в).
При развитии зигоспоры происходит слияние двух внешне неразличимых клеток мицелия, а при развитии ооспоры – слияние двух внешне различных половых клеток.
Ооспоры и зигоспоры имеют толстую оболочку, содержат много запасных питательных веществ и способны долго сохраняться в неблагоприятных условиях.
Большинство грибов может размножаться бесполым и половым путем, такие грибы называют совершенными. Некоторые грибы не способны к половому размножению, их называют несовершенными. Особенности способов размножения и строения органов размножения используют при распознавании грибов. Эти особенности лежат в основе их классификации.
Дата добавления: 2014-12-15; просмотров: 14809;