ГЛАВА 5. НЕРВНАЯ СИСТЕМА 4 страница
Лимфатические сосуды головы и шеи собираются в правый и левый яремные лимфатические стволы, trunci jugulares dexter et sinister: truncus jugularis dexter впадает в ductus lymphaticus dexter, truneus jugularis sinister — в ductus thoracicus.
В области головы и шеи (рис. 291) различают следующие основные группы лимфатических узлов.
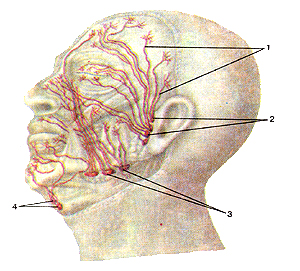
Рисунок 291. Лимфатические сосуды и узлы головы. Вид слева. 1 - лимфатические сосуды; 2 - поверхностные околоушные лимфатические узлы; 3 - поднижнечелюстные лимфатические узлы; 4 - подбородочнне лимфатические узлы.
1. Затылочные лимфатические узлы, nodi lymphatici occipilales, залегают в подкожной клетчатке на уровне верхней выйной линии.
2. Заушные лимфатические узлы, nodi lymphatici retroauriculares, позади ушной раковины.
3. Поднижнечелюстные лимфатические узлы, nodi lymphatici submandibulares в trigonum submandibulare (часть их залегает в толще поднижнечелюстной слюнной железы).
4. Подподбородочные лимфатические узлы, nodi lymphatici submentales, выше тела подъязычной кости, на передней поверхности mm. mylohyoidei.
5. Нижнечелюстные лимфатические узлы, nodi lymphatici mandibulares.
6. Лимфатические узлы околоушной железы, nodi lymphatici parotidei, — в окружности и толще околоушной железы; различают поверхностные и глубокие, nodi lymphatici parotidei superficiales et profundi.
7. Щечные лимфатические узлы, nodi lymphatici buccales.
8. Язычные лимфатические узлы, nodi lymphatici linguales, по бокам корня языка.
9. Поверхностные шейные лимфатические узлы, nodi lymphatici cervicales superficiales, no ходу наружной яремной вены и позади m.. sternocleidomastoideus.
10. Глубокие шейные лимфатические узлы, nodi lymphatici cervicales profundi, делятся на двубрюшно-яремные, nodi lymphatici jugulodigastrici, залегающие по ходу магистральных сосудов от основания черепа до уровня деления общей сонной артерии, и яремно-лопаточно-подьязычные, nodi lymphatici juguloomohyoidei, располагающиеся книзу и кзади от ключицы.
11. Заглоточные лимфатические узлы, nodi lymphatici retropharyngei, располагающиеся по боковой поверхности и несколько позади глотки.
Поверхностные лимфатические сосуды головы начинаются из лимфатических сетей кожи и делятся на две группы: переднюю и заднюю. Направление крупных лимфатических сосудов соответствует ходу кровеносных сосудов. Задняя группа поверхностных лимфатических сосудов головы собирает лимфу от затылочной области, от задней половины теменной и височной областей, от ушной раковины, наружного слухового прохода и барабанной перепонки. Лимфатические сосуды затылочной области вступают в затылочные лимфатические узлы, nodi lymphatici occipitales (числом 2-3). Лимфатические сосуды теменной и височной областей и ушной раковины направляются к заушным лимфатическим узлам, nodi lymphatici retroauriculares (числом 3-4). Лимфатические сосуды от барабанной перепонки, наружного слухового прохода и части ушной раковины идут к поверхностным и глубоким лимфатическим узлам околоушной железы, nodi lymphatici parotidei superficiales et profundi. Выносящие лимфатические сосуды этих узлов (nodi occipitales, retroauriculares, parotidei) — впадают преимущественно в поверхностные шейные лимфатические узлы, nodi lymphatici cervicales superficiales; часть сосудов направляется в глубокие шейные лимфатические узлы, nodi lymphatici cervicales profundi. Передняя группа поверхностных лимфатических сосудов головы начинается в лимфатических сетях кожи лба, наружного отдела верхнего и нижнего века, передних отделов теменной и височной областей и передней поверхности ушной раковины. Лимфатические сосуды указанных областей направляются к nodi lymphatici parotidei superficiales, располагающимся впереди ушной раковины и у верхнего края околоушной железы. Выносящие сосуды этих узлов вступают в толще околоушной железы в глубокие лимфатические узлы околоушной железы, nodi lymphatici parotidei profundi, выносящие сосуды которых на уровне угла нижней челюсти впадают в глубокие шейные лимфатические узлы, nodi lymphatici cervicales profundi. Лимфатические сосуды из кожных сетей внутренних половин верхнего и нижнего века, надпереносья, носа, щеки и верхней и нижней губ, а также глубокие сосуды от мышц, костей, слизистой оболочки преддверия рта и носа, конъюнктивы направляются соответственно ходу vasa facialia в подчелюстной треугольник, где вступают в поднижнечелюстные лимфатические узлы, nodi lymphatici suhmandibulares. числом от 6 до 10. Часть указанных лимфатических сосудов прерывается в щечных лимфатических узлах, nodi lymphatici huccales, находящихся на наружной поверхности m. buccinator. Лимфатические сосуды от нижней губы и подбородка направляются к подподбородочным лимфатическим узлам, nodi lymphatici submentales. которые находятся над телом подъязычной кости; они принимают также лимфу от лимфатических сосудов кончика языка. Глубокие лимфатические сосуды от твердого и мягкого неба, носоглотки и полости носа, крылонебной и подвисочной ямок направляются к глубоким лицевым лимфатическим узлам и к nodi lymphatici parotidei. Лимфатические сосуды языка делятся на поверхностные, которые начинаются из сети лимфатических сосудов слизистой оболочки, и на глубокие, сопровождающие кровеносные сосуды. Обе группы сосудов вступают в язычные лимфатические узлы, nodi lymphatici linguales. Отводящие сосуды языка направляются к nodi cervicales profundi, submandibulares et submentales. Лимфатические сосуды от верхнего и нижнего века, конъюнктивы и глазницы направляются в соответствующие регионарные узлы. Глазное яблоко лимфатических сосудов не имеет, но содержит лимфатические пространства. К ним относятся spatia zonularia (так называется лимфатическое пространство между расходящимися волокнами связки, подвешивающей хрусталик), передняя и задняя камеры глаза и щели между оболочками. Отток жидкости из передней и задней камеры и spatia zonularia осуществляется через spatia anguli iridocornealis (так называются микроскопической величины лимфатические щели между пучками гребенчатой связки радужно-роговичного угла глазного яблока) в венозную пазуху склеры, sinus venosus sclerae, а оттуда — в венозную систему.
Поверхностные лимфатические сосуды шеи направляются к v. jugularis externa, в окружности которой они соединяются между собой и вступают в nodi lymphatici cervicales superficiales, числом до 4-5. Глубокие лимфатические сосуды шеи собирают лимфу от внутренних органов шеи — глотки, гортани, трахеи и шейной части пищевода, щитовидной железы и мышц шеи — и направляются к сосудисто-нервному пучку шеи, где вступают в двубрюшно-яремный лимфатический узел и в глубокие шейные лимфатические узлы, nodi lymphatici jugulodigastrici et nodi lymphatici cervicales profundi. Лимфатические сосуды боковых долей щитовидной железы вливаются в двубрюшно-яремные лимфатические узлы; лимфати-ческие сосуды перешейка щитовидной железы предварительно прерываются в предгортанных лимфатических узлах, которые, числом 2-3, лежат над верхним краем перешейка, и в трахеальных лимфатических узлах, которые находятся ниже перешейка на боковой поверхности трахеи. Указанные узлы принимают также ряд лимфатических сосудов от гортани. По ходу лимфатических сосудов глотки находятся заглоточные лимфатические узлы, nodi lymphatici retropharyngei, располагающиеся на заднебоковой поверхности глотки. Выносящие сосуды перечисленных узлов вливаются в глубокие шейные лимфатические узлы, nodi lymphatici cervicales profundi. Последние вместе с подходящими сюда лимфатическими сосудами образуют яремное лимфатическое сплетение; их сосуды направляются к яремно-лопаточно-подъязычному лимфатическому узлу и глубоким шейным лимфатическим узлам, nodi lymphatici juguloomohyoidei et cervicales profundi, которые собирают всю лимфу от головы и шеи; они залегают, числом 10-15, от уровня деления сонной артерии до ключицы, располагаясь на передней поверхности лестничных мышц. Лимфа оттекает от них в правый лимфатический проток, ductus lymphaticus dexter, справа и в грудной проток, ductus thoracicus, слева. Во все перечисленные узлы соответственно вливаются также лимфатические сосуды нижнего отдела глотки, шейного отдела пищевода и трахеи.
В области верхней конечности (рис. 292) различают следующие лимфатические узлы.
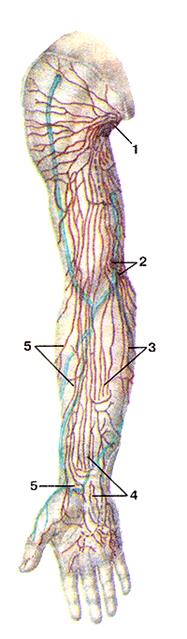
Рисунок 292. Лимфатические сосуды и узлы верхней конечности, правой. Вид спереди.
1 - подмышечные лимфатические узлы; 2 - локтевые лимфатические узлы; 3 - медиальные лимфатические сосуды; 4 - промежуточные (средние) лимфатические сосуды; 5 - латераль-ные лимфатические сосуды.
1. Подмышечные лимфатические узлы, nodi lymphatici axillares, числом 15-20, залегают в подмышечной ямке. Они являются регионарными узлами верхней конечности и области пояса верхней конечности. Часть подмышечных узлов располагается поверхностно, в слое жировой клетчатки, остальные — в глубине подмышечной ямки, в окружности кровеносных сосудов. В зависимости от положения в них различают: верхушечные, центральные, латеральные и грудные лимфатические узлы, nodi lymphatici apicales, centrales, laterales. pectorales. По топографии и связям с лимфатическими сосудами определенных областей лимфатические узлы верхней конечности делят также на три группы: передние, нижние и наружные. Передняя группа узлов (nodi lymphatici pectorales) располагается на наружной поверхности m. serratus anterior по ходу a. thoracia lateralis и принимает лимфу от поверхностных сосудов верхнего отдела передней брюшной стенки, переднебоковых отделов грудной клетки и молочной железы. Нижняя группа узлов (nodi lymphatici centrales et subscapulares) находится в заднем отделе подмышечной ямки. Эта группа узлов принимает лимфатические сосуды плеча и задней поверхности грудной клетки. Наружная группа узлов (nodi lymphatici laterales) залегает на наружной стенке подмышечной ямки и принимает лимфатические сосуды верхней конечности.
2. Плечевые лимфатические узлы располагаются по ходу плечевой артерии.
3. Локтевые лимфатические узлы, nodi lymphatici cubitales, располагаются преимущественно в глубоких отделах локтевой ямки в окружности кровеносных сосудов. Часть этих узлов (1-3) залегает поверхностно над медиальным надмыщелком плечевой кости.
4. Лимфатические узлы предплечья, числом 1-2, находятся в верхней трети предплечья по ходу локтевой артерии. Лимфатические сосуды верхней конечности делятся на поверхностные и глубокие лимфатические сосуды.
Поверхностные лимфатические сосуды верхней конечности залегают в поверхностных слоях подкожной клетчатки. Они начинаются из лимфатических сетей тыльной и ладонной поверхностей кисти, образуя две группы крупных лимфатических сосудов: внутреннюю по ходу v. basilica и наружную по ходу v. cephalica. Крупные стволы поверхностных сосудов, числом 8-10, по ходу принимают мелкие лимфатические сосуды смежных областей. Внут-ренняя группа поверхностных лимфатических сосудов верхней конечности, следуя по ходу v. basilica, достигает локтевой ямки. Здесь один — два сосуда вступают в локтевые лимфатические узлы, nodi lymphatici cubitales, выносящие сосуды которых идут вместе с веной под плечевую фасцию к глубоким лимфатическим сосудам плеча. Остальные лимфатические сосуды этой группы следуют в подкожной клетчатке по внутренней поверхности плеча и достигают nodi lymphatici axillares. Наружная группа поверхностных лимфатических сосудов верхней конечности направляется вместе с v. cephalica и, достигнув верхней трети плеча, проникает с ней в глубину, в подмышечную ямку, где также достигает nodi lymphatici axillares.
Глубокие лимфатические сосуды верхней конечности собирают лимфу от мышц, костей и суставов. Лимфатические сосуды пальцев идут по их боковой поверхности по ходу артерий. На кисти эти сосуды, анастомозируя между собой, образуют ладонное лимфатическое сплетение, которое соответствует артериальной дуге. Отводящие лимфатические сосуды этого сплетения идут на предплечье, располагаясь по ходу a. radialis и a. ulnaris. Лимфатический сосуд, идущий по ходу v. ulnaris, прерывается в верхней трети предплечья в лимфатических узлах предплечья, куда вливается также лимфатический сосуд, собирающий лимфу от тыла предплечья и сопровождающий заднюю межкостную артерию. Лимфатический сосуд, сопровождающий лучевую и локтевую артерии, достигнув локтевой ямки, вступает в nodi lymphatici cubitales. Выносящие сосуды этих узлов образуют одиночный лимфатический сосуд, который направляется на плечо по ходу а. brachialis. На границе нижней и средней трети плеча указан-ный сосуд вступает в лимфатический узел плеча, из которого выходят два выносящих сосуда. Поднимаясь кверху по наружной и внутренней поверхностям плечевой артерии, они достигают подмышечной ямки, где вступают в наружную группу подмышечных лимфатических узлов. Лимфатические сосуды верхнего отдела передней брюшной стенки (поверхностные) кнаружи и кверху от пупка прерываются в надчревном лимфатическом узле, nodulus lymphalicus epigastricus, и, следуя по боковой поверхности грудной клетки, достигают подмышечной ямки, где вступают в переднюю группу nodi lymphatici axillares.
ГЛАВА 5. НЕРВНАЯ СИСТЕМА
Нервная система человека (рис. 293) подразделяется на центральную (головной и спинной мозг) ипериферическую(нервные корешки, узлы, сплетения, черепные и спинномозговые нервы).
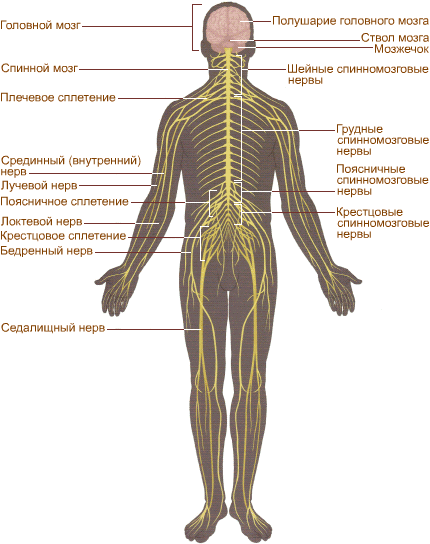
Рисунок 293. Схема нервной системы человека
На рисунке схематично показана нервная система человека. Она играет главную роль в согласованной деятельности организма. Особый комплекс периферических нервов и нервных узлов, иннервирующих сердце, легкие, пищеварительный тракт и другие внутренние органы, сосуды и ткани, и есть вегетативная нервная система. Ее работа, как правило, не зависит от волевых усилий человека, и в норме мы не ощущаем раздражения вегетативных узлов и нервов. Нервные узлы симпатического отдела вегетативной нервной системы образуют симпатические нервные стволы, расположенные около спинного мозга, а нервные узлы другого отдела — парасимпатического – лежат во внутренних органах или около них.
Для слаженной деятельности различных частей такой сложной системы, как организм человека, необходимо координирующее устройство соответственной сложности. И в самом деле, нервная система, интегрирующая деятельность всех частей тела, является, несомненно, самой сложной из всех систем органов. Мышцы и железы животного или человека носят общее название эффекторов; глаза, уши и другие органы чувств называются рецепторами. Нервная система, состоящая из головного мозга, спинного мозга и проводящих путей, соединяет рецепторы с эффекторами и передает импульсы, или «сообщения», от первых ко вторым. Она способна делать это таким образом, что при раздражении того или иного рецептора должным образом реагирует надлежащий эффектор. Основными функциями нервной системы являются проведение импульсов и интеграция деятельности различных систем организма. Координирующие функции нервной системы, эндокринная регуляция и собственные регуляторные механизмы внутриклеточных ферментных систем (торможение и стимуляция активности ферментов, индукция и репрессия их синтеза) — все это факторы, способствующие гомеостазу, т.е. поддержанию постоянства внутренней среды организма.
Центральная нервная система (ЦНС) — это совокупность нервных образований спинного и головного мозга, обеспечивающих восприятие, обработку, передачу, хранение и воспроизведе-ние информации с целью адекватного взаимодействия организма и изменений окружающей среды, координации оптимальной работы органов, их систем и организма в целом.
Каждая из этих структур имеет морфологическую и функциональную специфику. Но, наряду с этим, у всех структур нервной системы есть ряд общих свойств и функций, к которым относятся: нейронное строение, электрическая и химическая синаптическая связь между нейронами, образование локальных сетей из нейронов, реализующих специфическую функцию, множественность прямых и обратных связей между структурами, способность нейронов всех структур к восприятию, обработке, передаче и хранению информации, преобладание числа входов для ввода информации над числом выходов, способность к параллельной обработке информации, способность к саморегуляции, функционирование на основе рефлекторного доминантного принципа.
Головной мозг является важнейшим отделом ЦНС, в нем различают стволовую часть и конечный мозг, включающего подкорковые или базальные ганглии и большие полушария.
Основные части головного мозга выделяются уже к 3-му месяцу эмбрионального развития, а к 5-му месяцу эмбриогенеза уже хорошо заметны основные борозды больших полушарий.
К моменту рождения общая масса головного мозга составляет около 400 г., причем у девочек он несколько меньше (388 и 391 у девочек и мальчиков соответственно). По отношению к массе тела мозг у новорожденного значительно больше, чем у взрослого. Так, если у новорожденного он составляет 1/8 массы тела, то у взрослого — 1/40. Наиболее интенсивно головной мозг человека развивается в первые два года постнатального развития. Затем темпы его роста снижаются, но продолжают оставаться высокими до 6-7 лет, к этому моменту масса мозга достигает уже 4/5 массы взрослого мозга. Окончательное созревание головного мозга заканчивается только к 17-20 годам. К этому возрасту масса мозга увеличивается по сравнению с новорожденными в 4-5 раз и составляет в среднем у мужчин 1400 г, а у женщин — 1260 г. Следует отметить, что абсолютная масса мозга не определяет непосредственно умственные способности человека.
Изменения размеров, формы и массы мозга сопровождается изменением его внутренней структуры. Усложняется строение нейронов, форма межнейронных связей, становится четко разграниченным белое и серое вещество, формируются различные проводящие пути головного мозга.
Развития мозга, как и других систем, идет гетерохронно. Раньше других созревают те структуры, от которых зависит нормальная жизнедеятельность организма на данном возраст-ном этапе. Функциональной полноценности достигают вначале стволовые, подкорковые и корковые структуры, регулирующие вегетативные функции организма. Эти отделы по своему развитию приближаются к мозгу взрослого человека уже к 2-4 годам постнатального периода.
Продолговатый мозг, мост, средний мозг, промежуточный мозг и мозжечок относятся к стволу мозга. В филогенетическом отношении это наиболее древние нервные структуры и поэтому их функции тесно связаны с регуляцией примитивных функциональных процессов.
В процессе онтогенеза созревание структур стволовой части головного мозга наиболее интенсивно происходит в первые два года жизни. Окончательное формирование этих структур, особенно промежуточного мозга, завершается только в 13-16 лет, когда заканчивается половое развитие подростков. Многие особенности низшей и высшей нервной деятельности у детей подросткового возраста объясняются функциональными свойствами промежуточного мозга и некоторых других подкорковых структур.
Наиболее молодым в филогенетическом отношении является конечный мозг. В его состав входят большие полушария и расположенные под ними скопления серого вещества в виде подкорковых или базальных ганглиев.
Большие полушария осуществляют регуляцию высших нервных функций, лежащих в основе всех психических процессов человека. Правое и левое полушарие тесно связаны между собой с помощью огромного количества нервных волокон, образующих мозолистое тело. Многие нервные процессы, выходящие из какой-либо точки одного полушария, проецируются в сим-метричную точку другого полушария. Таким образом, в нервной деятельности полушарий конечного мозга проявляется свойство билатеральной симметрии.
Существует предположение, что в процессе онтогенеза развитие парной деятельности полушарий идет от неустойчивой симметрии к неустойчивой асимметрии, и наконец, к устойчивой функциональной асимметрии. Это подтверждается развитием бимануальных действий человека, то есть особенностей двигательной деятельности левой и правой рук. Пока-зано, что к праворукости дети переходят с 2-4 лет. В этом возрасте правши составляют 38%, а к 4-6 годам — 75%. Иначе говоря, в значительной степени моторная асимметрия зависит от условий воспитания ребенка, но вместе с тем существует и наследственная предраспо-ложенность.
Темпы созревания левого и правого полушарий имеют половые особенности. Левое полу-шарие у девочек развивается быстрее, что свидетельствует о более раннем созревании доминантного полушария. Данный факт косвенно подтверждается также более быстрым развитием у девочек речи и некоторых показателей психомоторики.
Высшим центром регуляции и управления всей деятельностью организма, начиная от самых примитивных физиологических отправлений и кончая сложнейшими психическими процес-сами у человека, является кора головного мозга. Активное формирование полушарий мозга начинается с 12-й недели эмбриогенеза и интенсивно продолжается в первые годы постнаталь-ного развития, особенно до 2 лет. Клеточное строение, форма и расположение борозд и извилин приближается к взрослому мозгу в 7 лет. А в лобных долях это различие сглаживается только к 12 годам. Существует прямая зависимость между морфофункциональным созреванием лобных долей больших полушарий и формированием психических функций у детей. Окончательное созревание больших полушарий и коры мозга завершается к 20-22 годам.
Морфологический анализ процессов созревания КГМ ребенка на клеточном уровне свидетельствует о постоянном увеличении размеров высших первичных, вторичных и третич-ных зон КГМ в процессе постэмбрионального развития: чем больше возраст ребенка, тем большие размеры занимают эти корковые зоны, и тем сложнее становится психическая деятельность.
Таким образом, в процессе постнатального развития происходит совершенствование морфо-логического строения КГМ, а параллельно этому и совершенствование высшей нервной деятельности ребенка и его психических процессов. Например, поля двигательного центра речи достигают функциональной полноценности только к 7 годам, к этому возрасту они увеличи-ваются на 64-73% в сравнении с мозгом новорожденного. То же можно сказать и о корковых зонах, ответственных за интеграцию слуховых и зрительных раздражителей, что имеет большое значение в формировании речи.
Важные данные о функциональной зрелости коры и подкорковых образований мозга и участия их в восприятии афферентных сигналов в разные возрастные периоды получены при использовании электрофизиологических методов. Анализ имеющихся в литературе данных о характере фоновой и вызванной электрической активности мозга человека на разных этапах онтогенеза рассматривается в связи с проблемой созревания высших отделов центральной нервной системы.
В раннем постнатальном периоде наиболее функционально зрелыми являются мезодиэнце-фалические структуры мозга, определяющие ЭЭГ-картину глубокого сна и реакцию возбуждения у новорожденного ребенка. Ряд факторов свидетельствует, что кора больших полушарий начинает функционировать уже с момента рождения ребенка. Нервные элементы коры больших полушарий новорожденного способны продуцировать кратковременную ритмическую электрическую активность. Это выражается в виде: 1) наличия групп синхронизи-рованных ЭЭГ-колебаний в затылочных областях мозга в переходном от бодрствования ко сну состоянии; 2) реакции усвоения ритма световых мельканий, наблюдаемой с первых часов жизни ребенка; 3) наличия ритмического сенсорного разряда, регистрируемого в затылочной области коры при значительной стимуляции. Отмечается, что кора больших полушарий новорожденных вовлекается в реакцию при афферентных воздействиях. В этом случае наблюдаются как генерализованные изменения электрической активности, обусловленные возбуждением подкорковых неспецифических структур мозга, так и локальные вызванные ответы, свидетельствующие о поступлении сигнала в кору больших полушарий по специфическому афферентному пути. Наличие ответных специфических и неспецифических реакций на афферентное раздражение означает функционирование восходящих ретикулярных и таламокортикальных связей, посылающих сенсорную информацию в кору больших полушарий. Для оценки способности коры воспринимать приходящую информацию наибольший интерес представляет наличие с момента рождения ребенка начальной позитивности вызванного специфического ответа, свидетельствующее о непосредственном участии нейронов 3-го и 4-го слоев коры в приеме афферентного сигнала.
Подчеркивая функционирование коры больших полушарий в период новорожденности, следует иметь в виду и отличия в деятельности ее нервных элементов по сравнению со взрослыми. Одним из таких проявлений служит отсутствие синхронизированной ритмической активности во время бодрствования новорожденных. Устойчивая ритмика в ЭЭГ бодрствую-щих детей регистрируется только с 2-3 месяцев постнатальной жизни. Появление организован-ной ритмики в состоянии спокойного бодрствования отражает важный этап в созревании коры мозга ребенка. В этом возрасте исчезают архаические рефлексы, развивается оптомоторная пространственная координация.
В течение первого года жизни формируется строго ритмическая электрическая активность частотой 5 Гц с фокусом в затылочной области коры, которая может рассматриваться как аналог альфа-ритма взрослого человека. С возрастом отмечается прогрессивное учащение альфа-ритма, появление и стабилизация его в центральных областях коры. Формирование основного ритма электрической активности, отражающее морфофункциональное созревание нейронного аппарата коры больших полушарий, заканчивается к 16-18-летнему возрасту. Созревание нервных элементов коры больших полушарий проявляется также в эволюции специфически вызванных потенциалов. В процессе индивидуального развития ребенка отмечается укорочение временных параметров ответа, усложнение его компонентного состава и появление вызванных потенциалов в ассоциативных областях коры. Включения ассоциа-тивных зон в прием и переработку качественно специфической информации, вероятно, опре-деляет возможность синтеза интегрального образа раздражителей разного информационного значения.
Параллельно с изменениями ЭЭГ, обусловленными функциональным созреванием коры больших полушарий, отмечается уменьшение выраженности подкорковых знаков в ЭЭГ (тета - волн, билатеральных пароксизмальных разрядов, усиленных неспецифических ответов). Ослаб-ление подкорковых знаков в ЭЭГ с возрастом можно объяснить усилением тормозных влияний созревающей коры на подкорковые структуры.
Таким образом, выявляются определенные этапы функционального созревания коры и подкорковых структур мозга, специфика их взаимных влияний в различные возрастные периоды.
Нервная ткань состоит из двух разновидностей клеток: нервных (нейронов) и глиальных. Глиальные клетки вплотную прилегают к нейрону, выполняя опорную, питательную, секретор-ную и защитную функции.
Вся нервная система построена на нервной ткани. Нервная ткань состоит из нервных клеток (нейронов) и связанных с ними анатомически и функционально вспомогательных клеток нейроглии. Нейроны выполняют специфические функции, являясь структурно-функциональной единицей нервной системы. Нейроглия обеспечивает существование и специфические функции нейронов, выполняет опорную, трофическую (питательную), разграничительную и защитную функции.
Нейрон (нейроцит) получает, перерабатывает, проводит и передает информацию, закоди-рованную в виде электрических или химических сигналов (нервных импульсов).
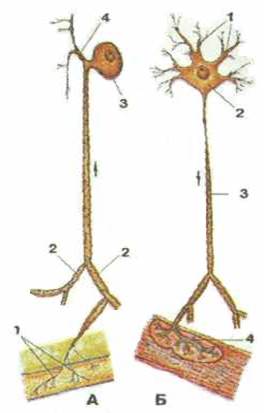
Рисунок 294. Нервные клетки. А — чувствительный нейрон. Б — двигательный нейрон. Стрелки показывают направление следования нервных импульсов. А: 1 - чувствительные нервные окончания; 2 - дендриты; 3 - тело нервной клетки; 4 - аксон. Б: 1 - дендриты; 1 - тело нервной клетки; 3 - аксон; 4 - двигательное нервное окончание (нервно-мышечная бляшка).
Каждый нейрон имеет тело, отростки и их окончания (рис. 294). Снаружи нервная клетка окружена оболочкой (цитолеммой), способной проводить возбуждение, а также обеспечивать обмен веществ между клеткой и окружающей их средой. Тело нервной клетки содержит ядро и окружающую его цитоплазму (перикарион). Цитоплазма нейронов богата органеллами (субклеточными образованиями, выполняющими ту или иную функцию). Диаметр тел нейро-нов варьирует от 4-5 до 135 мкм. Форма тел нервных клеток тоже различная — от округлой, овоидной до пирамидальной. От тела нервной клетки отходят различной длины тонкие отростки двух типов. Один или несколько древовидно ветвящихся отростков, по которым нервный импульс приносится к телу нейрона, называют дендритом. У большинства клеток их длина составляет около 0,2 мкм. Единственный, обычно длинный отросток, по которому нервный импульс направляется от тела нервной клетки — это аксон, или нейрит.
По количеству отростковнейроны подразделяются на униполярные, би- и мульти-полярные клетки. Униполярные (одноотростчатые) нейроны имеют лишь один отросток. У человека такие нейроны встречаются лишь на ранних стадиях внутриутробного развития. Биполярные (двухотростчатые) нейроны имеют один аксон и один дендрит. Их разно-видностью являются псевдоуниполярные (ложноуниполярные) нейроны. Аксон и дендрит этих клеток начинаются от общего выроста тела и в последущем Т-образно делятся. Мультиполяр-ные (многоотросчатые) нейроны имеют один аксон и много дендритов, они составляют большинство в нервной системе человека. Нервные клетки динамически поляризованы, т.е. способны проводить нервный импульс только в одном направлении — от дендритов к аксону.
Дата добавления: 2014-12-12; просмотров: 784;