Центральные механизмы, участвующие в регуляции кровяного давления барорецепторами
Мы не можем шодробно описывать здесь все рефлекторные механизмы регуляции -кровяного давления. Укажем лишь, что давление в артериях определяется количеством -находящейся в лих крови, т. е. соотношением между притоком и оттоком. Количество крови, :по-стулающсй в артерии в единицу времени, определяется минутлы-м объемом, а скорость оттока зависит главлым образом от толуса артериол, регулирующего сопротивление оттоку. Прессорные центры продолговатого мозга генерируют нервные импульсы, которые ведут к сокра-
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 157
щению артериол и .повышению частоты сердечных сокращений. Это сосудосуживающий и ускоряющий центры, возбуждение которых вызывает сужение сосудов и ускорение сердечного ритма через симпатические нервы. Повышая приток крови в артерии (минутный объем) и уменьшая отток (повышение периферического сопротивления), эти прессорные центры повышают артериальное давление.
Наоборот, депрессорные центры имеют тенденцию снижать артериальное давление путем замедления сердечного ритма (минутный объем) и снижения степени сокращения артериол. Тормозной центр сердца оказывает свое влияние через (парасимпатический) блуждающий нерв. Расширение сосудов в значительной степени является следствием снижения активности сосудосуживающего центра.
Депрессорный характер зависит от того влияния, которое оказывают импульсы с барорецепторов на центры регуляции кровяного давления в продолговатом мозге. Под действием этих импульсов возбудимость депрессор-ных центров повышается, а прессорных— снижается. Мы уже подчеркивали, что рефлексы с барорецепторов имеют чисто депрессорный характер. Импульсы от барорецепторов не вызывают повышения артериального давления. Однако тот факт, что давление может резко повыситься рефлекторным путем 'при снижении или выключении активности барорецепторов, дал основание для неточных высказываний, под влиянием которых могло сложиться впечатление, что это повышение зависит от барорецепторных рефлексов. Фактически повышение давления зависит от устранения двпрессорного действия барорецепторов, что позволяет различным факторам, повышающим кровяное давление, действовать без ограничения.
Таким образом, мы видим, что колебания артериального давления /ограничиваются благодаря изменению разрядов ;барорецепторов. Падение давления ограничивается снижением депреосорных влияний, а повышение— их увеличением. По этой причине нервы, идущие от каротидного синуса и дуги аорты, получили название буферных.
ГЛАВА V
ВЛИЯНИЕ РЕФЛЕКСОВ С БАРОРЕЦЕПТОРОВ НА ГИПОТАЛАМУС
Дальнейшие исследования .показали, что рефлексы с барорецепторов влияют не только на центры .продолговатого мозга, ио и на гипоталамус. Регистрируя электрическую активность гипоталамуса, можно показать, что при искусственном снижении кровяного давления в симпатическом отделе гипоталамуса наблюдается активация. Поскольку мы уже знаем, что падение давления вызывает снижение или исчезновение разрядов в баро-рецепторах, ясно, что эти разряды каким-то образом подавляют активность симпатического отдела гипоталамуса. Следовательно, влияние импульсов с барорецепторов на симпатический отдел гипоталамуса аналогично их действию на симпатические центры продолговатого мозга. Если, введя в 'гипоталамус небольшое количество стрихнина, вызвать описанные выше стрихнинные пики, то возбуждение гипоталамуса при падении кровяного давления выразится в форме повышения частоты разрядов. Если, напротив, вместо снижения кровяного давления вызвать в эксперименте его повышение, активность гипоталамуса подавляется.
Установлено, что эти глубокие изменения возбудимости гипоталамуса оказывают существенное влияние на кору головного мозга. В качестве индикатора и в этом случае использовалась запись электрической активности. Было обнаружено, что при снижении кровяного давления повышенная активность регистрируется не только н гипоталамусе, но и в ,коре мозга; при повышении кровяного давления наблюдается обратная картина [674].
Эти сдвиги зависят от влияния рефлексов с барорецепторов на гипоталамус, а через гипоталамо-корти-кальные разряды и на кору головного мозга. Это было доказано следующими наблюдениями и экспериментами.
Если изменения активности гипоталамуса и коры обусловлены рефлексами с 'барорецепторов, то эти изменения не должны зависеть от причины, вызвавшей --. изменение кровяного давления.
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 159
Интенсивность разрядов 'барорецепторов определяется самим изменением давления, а не характером фармакологического препарата, вызвавшего это изменение. Эксперименты показали, что эффект возбуждения гипо-таламо-кортикальной системы возникает при снижении кровяного давления любыми химическими или механическими оредствами. Фармакологические препараты, резко различные по своей химической природе, такие, как гистамин или ацетилхолин, дают совершенно одинаковый физиологический эффект. Еще 'более убедительные доказательства того, что эти вещества действуют на гипоталамус и кору через рефлексы каротидяо-аорталь-ных барорецепторов, были получены в опытах с перерезкой барорецепторных нервов. После такой перерезки снижение кровяного давления введением ацетилхолина не вызывает возбуждения гипоталамо^ортикальной системы— электрическая активность коры остается неизменной. Не изменяется эта активность и при повышении кровяного давления введением адреналина. Отсюда мож*но сделать вывод, что изменения кровяного давле-| ния действуют на гипоталамус и кору головного мозга через рефлексы с каротидяо-аорталыных барорецепто ров [310].
Влияние рефлексов с барорецепторов на симпатическую систему
Установив, что сдвиги кровяного давления вызывают изменения активности гипоталамуса и коры мозга через рефлексы с барорецепторов,' мы, естественно, задаем себе вопрос, как меняется вегетативная активность на уровне гипоталамуса под влиянием этих воздействий. Для ответа на этот вопрос был проведен следующий эксперимент. Под поверхностным наркозом кошке вводили электрод в задний гипоталамус и определяли реакцию на электрическое раздражение (тестирующее 'раздражение) путем регистрации кровяного давления сердечного ритма и сокращения третьего века. Затем кРовяное давление изменяли и вновь применяли то же Раздражение.
I 60,
ГЛАВА V
В тервой серии опытов снижение кровяного давления вызывали введением ацетилхолина, мехолила или ги-стамина. Затем, по мере восстановления нормального давления вновь применяли тестирующее раздражение. На фиг. 21 .показаны результаты контрольного эксперимента, в котором тестирующее раздражение производили на фоне нормального кровяного давления, отмечено небольшое (повышение кровяного давления и умеренное

Фиг. 21. Повышенная симпатическая реактивность при раздражении заднего отдела гипоталамуса на фоне понижения кровяного давления, вызванного ацетилхолином [310].
/ — внутривенное введение ацетилхолина (10 мкг); 2— раздражение заднего отдела правого гипоталамуса (0,6 в. 99 имп/сек, 0,4 мсек, 2 сек\; 3 — комбинация / и 2. I ~ сокращение третьего века; // — кровяное давление.
сокращение третьего века, имеющего симпатическую иннервацию, т. е. типичная слабо выраженная симпатическая реакция. В противоположность этому применение того же тестирующего раздражения после снижения кровяного давления, вызванного ацетилхолином, приводит ,к резкому сокращению третьего века и быстрому повышению кровяного давления (фиг. 21). Этот эксперимент можно воспроизвести много раз на одном и том же животном. Повышенная реактивность симпатической системы документировалась одним, двумя либо всеми тремя использованными индикаторами (кровяное давление, сердечный ритм и сокращение третьего века) в зависимости от таких факторов, 1ка>к вид раздражителя и общая чувствительность препарата [358].
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 161
В другой 'Серии опытов было показано, что повышение реактивности симпатического отдела гипоталамуса под влиянием падения кровяного давления зависит от целости афферентных нервов барорецепторов. Так, на
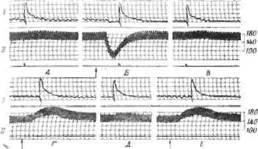
Фиг. 22. Отсутствие влияния изменений кровяного давления на
реактивность симпатической системы после денервации сино-аор-
тальной зоны у кошки [319].
/ — сокращение третьего века; Л —кровяное давление. Л, В и Д. Контрольные раздражения заднего отдела левого гипоталамуса (1,5 а, 99 имп/сек, 1.R мсек, 1 сек). Б. Кровяное давление снижено ацстилхолином (0,0019 мг, внутривенно), после чего произведено раздражение гипоталамуса. Г, Е. Введение 0,4 мкг норадреналина и раздражение гипоталамуса. Характерно, что интенсивность вызванного раздражением гипоталамуса сокращения, сохранившего иннервацию третьего века, остается во время медикаментозной гипотонии и гипертонии таким же, как и при нормальном давлении, в отличие от повышенной и ослабленной реакции у животного с сохраненной иннервацией сино-аортальной зоны.
фиг. 22 видно, что реакция третьего века на раздражение гипоталамуса остается неизменной во время вызванного ацетилхолином снижения давления, если каротидно-аор-тальная зона у животного денервирована. Из работ такого типа можно сделать вывод, что повышенная реактивность симпатического отдела гипоталамуса при различных формах экспериментально вызванной гипотонии обусловлена пониженной активностью барорецепторов [319].
Изменения реактивности гипоталамуса, "подобные тем, которые возникают при изменениях кровяного дав-
ГЛАВА V
ления, можно вызвать раздражителями, действующими непосредственно на гипоталамус. Самый простой опыт, иллюстрирующий это положение, приведен на фиг. 23. Раздражение левого заднего гипоталамуса в течение 20 сек вызывает незначительное повышение кровяного давления и некоторое ускорение сердечного ритма, изменения со стороны третьего века отсутствуют (фиг.. 23, А).
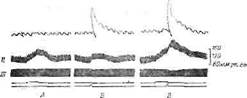
Фиг. 23. Пространственная симпатическая суммация у кошки [315].
/ — третье веко; // — кровяное давление; /// — пульс (снижение амплитуды указывает на учащение сердечного ритма). Л. Раздражение заднего отдела правого гипоталамуса (1 о, 158 имп/сек, 4 мсек. 20 сек). Б. Раздражение другой точки заднего отдела гипоталамуса (4,4 я. 180 имп/сек, 4 мсек, 1.5 сек). В. Комбинация обоих раздражений {А и Б), причем второй раздражитель применяется в течение последних 1,5 сек действия первого. Эффект суммацпи наблюдается по всем трем индикаторам.
Раздражение (правого заднего гипоталамуса в течение 1,5 сек приводит к незначительному повышению кровяного давления, сокращению третьего века и некоторому ускорению сердечного ритма (см. фиг. 23, Б). В третьем опыте оба раздражения сочетаются, причем более короткое (правый гипоталамус) начинается за 1 сек до прекращения более длительного (см. фиг. 23, В). Все три индикатора (третье веко, кровяное давление, сердечный ритм) свидетельствуют о более выраженной реакции, чем при действии .каждого из раздражений в отдельности. Особенно интересно, что хотя изолированное раздражение левого заднего гипоталамуса не вызывает сокращения третьего века, оно повышат реактивность гипоталамуса таким образом, что эффективность раздражения правого гипоталамуса возрастает. Такое облегчение, или настройка, может усилить реакцию
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 163
пюбого из использованных 'индикаторов или всех их вместе [315].
Значение этого явления для физиологии и патологии эмоций мы рассмотрим ниже. Здесь же отметим, что данный эксперимент служит моделью, иллюстрирующей тот факт, что симпатическая реактивность гипоталамуса может изменяться под действием центральных факторов. Таким образом, повышенная реактивность гипоталамуса может быть следствием возбуждения или облегчения его симпатического отдела независимо от того, вызвано оно непосредственно или рефлекторно. Даже подпороговые раздражители, т. е. те, которые не вызывают выраженной реакции со стороны избранного индикатора, способны обеспечить такое облегчение [324].
Влияние рефлексов с барорецепторов на парасимпатическую систему
На реактивность парасимпатического отдела гипоталамуса можно повлиять через рефлексы с барорецепторов путем введения гипертензивных веществ. Для этой цели использовали норадреналин. В основу исследования был положен тот же принцип, что и в опытах по изучению симпатической настройки. Парасимпатическую реакцию можно вызвать путем раздражения участков переднего гипоталамуса (парасимпатический отдел); для получения такой реакции при раздражении других отделов следует применять раздражение низкой частоты (3—8 в 1 сек.).
На фиг. 24, В показана парасимпатическая реакция на раздражение гипоталамуса с частотой 8 в 1 сек. Кровяное давление падает, а частота средечных сокращений снижается. Под > влиянием норадреналина (фиг. 24, А) кровяное давление повышается, и в результате рефлекторного влияния барорецепторов, вызванного повышением давления, вторично замедляется сердечный ритм. Если электрическое раздражение парасимпатического отдела гипоталамуса (фиг. 24, В) совпадает с парасимпатической реакцией, вызванной повышением кровяного давления (фиг. 24, А), парасимпатическая Реакция резко усиливается (фиг. 24,Б): происходит не
ГЛАВА V
только внезапное и выраженное замедление сердечного ритма, но и резкое падение кровяного давления.
Из этого эксперимента можно сделать вывод, что повышение 'Кровяного давления вызывает соответствующую (Настройку парасимпатических механизмов, так что парасимпатический эффект раздражения гипоталамуса

Фиг. 24. Пространственная парасимпатическая суммация у кошки под нембуталовым наркозом [315].
/ — кровяное давление; // — пульс. Л. Введение норадреналина (0,001 мг внутривенно). В. Раздражение заднего отдела гипоталамуса (3 в, 8 амщеек, 0,8 мсек, 7 сек). Б. Комбинация А и В. Одно только раздражение гипоталамуса (В) вызывает небольшую парасимпатическую реакцию, выражающуюся в незначительном снижении кровяного давления и столь же незначительном замедлении сердечного ритма. То же раздражение, примененное на фоне максимальной прессорной реакции, вызванной норадреналином, приводит к более резкому падению давления и более выраженному замедлению сердечного ритма.
усиливается [324]. Однако та^ая парасимпатическая настройка не ограничивается гипоталамусом, а распространяется и на другие 'парасимпатические центры. Подтверждением этому служит тот факт, что на фоне вызванной норадреналином парасимпатической настройки возрастает и эффективность рефлекторпо действующих парасимпатических раздражителей. Так, замедление сердечного ритма под влиянием низкочастотного раздражения седалищного нерва выражено значительно сильнее на фоне парасимпатической настройки, чем в контрольных условиях.
Как и в описанных выше экспериментах по изучению симпатической настройки, изменения кровяного давления действуют только через рефлексы с барорецепторов. Денервация баро1рецепторных зон исключает возможность парасимпатической настройки, возникающей в норме под влиянием гипертонии, аналогично тому, как она исключает описанную выше симпатическую настройку под влиянием гипотонии. Напомним, что разряд барорецепторов усиливается при повышении и ослаб-
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 165
чяется .при падении кровяного давления. Отсюда следует, что повышение афферентной импульсации бароре-цепторов приводит к парасимпатической настройке, а снижение афферентного разряда барорецепторов — к симпатической.
Суммируя эти данные, можно сказать, что вещества, влияющие на кровяное давление, изменяют реактивность гипоталамуса. Это обусловлено тесной связью между активностью барорецепторов и 'реактивностью гипоталамуса. Отклонение кровяного давления от нормального уровня вызывает изменение активности барорецепторов. В результате происходит настройка гипоталамуса, так что один из его отделов — симпатический или парасимпатический — становится более чувствительным, а другой соответственно менее чувствительным. (Такие реципрокные реакции противоположных систем будут освещены в следующем разделе.) Снижение кровяного давления приводит к симпатической настройке, облегчая таким образом восстановление нормального давления. Повышение давления, напротив, приводит к парасимпатической настройке, что также способствует восстановлению нормального уровня. Это гомеостатический механизм par excellence.
Дата добавления: 2014-12-10; просмотров: 1489;