РАЗРУШЕНИЯ, ДЕЛАЮЩИЕ ЖИВОТНЫХ БОЛЕЕ РУЧНЫМИ
В экспериментах Клювера и Быоси [123, 124, 544, 546], которые считаются классическими в литературе о поведении, превращали непокорных, диких обезьян в удивительно ручных, послушных животных путем двухстороннего удаления височных долей. После операции у этих обезьян не наблюдали ни признаков страха, ни реакции гнева, они не пытались сопротивляться, даже если с ними очень плохо обращались. Они, казалось, неспособны были отличать опасные предметы от полезных, т. е., по меткому определению Быоси и Клювера, они страдали «психической слепотой». Создавалось впечатление, что такие животные одержимы непреодолимым стремлением исследовать все доступные предметы:
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 111
они вновь и вновь обнюхивали их, брали в рот, бросали как несъедобные или причиняющие боль только для того, чтобы тут же снова повторить все сначала. Кроме того, им была свойственна общая гиперактивность и повышенная, но недифференцированная половая активность, выражавшаяся в мастурбации и попытках к совокуплению с особями любого пола. Пожалуй, первыми заметили подобные изменения поведения у амигдалэктомирован-ных обезьян Браун и Шефер [116] в 1888 г., но Быоси и Клювер привлекли особое внимание к этой проблеме своими глубокими исследованиями, результаты которых затем были подтверждены [4, 724, 943]. Хотя производимые ими разрушения мозга у обезьян обычно захватывали довольно обширную область, а не только миндалевидное ядро, в большинстве более поздних работ описанные нарушения получали при изолированном повреждении миндалевидного ядра [382]. Различные элементы синдрома Клювера—-Быоси были получены при разрушениях в различных частях лимбической системы обезьян [90].
Результат двухстороннего удаления миндалевидного ядра и лежащей над ним коры у кошек удивительно сходен с тем, что наблюдается у обезьян [95, 289, 541, 807, 808]. Отмечено заметное повышение болевого порога, гиперактивность, усиление реакций, выражающих удовольствие, гиперсексуальность и снижение страха и агрессивности. Шрайнер и Клинг [806а] получили такой результат в опытах на кошках и, кроме того, показали, что кошки, ставшие после амигдалэктомии ручными и гиперсексуальными, при последующем разрушении вент-ро-медиальных ядер гипоталамуса превращаются в злобных, гипосексуальных животных. Однако Морган и Косман [663] опубликовали несколько иные результаты. В течение 6 недель после второго этапа такой двухэтап-ной операции наблюдалась повышенная двигательная активность и голосовые реакции, но другие изменения поведения отсутствовали. Была отмечена трудность приручения этих кошек, но указано, что несколько животных, которые отличались «исключительно скверным характером», стали совершенно ручными без операции, после того как привыкли к новой обстановке. Единствен-
ГЛАВА IV
ное постоянное и определенное изменение, которое можно было приписать амигдалэктомии, была гиперфагия, которая оказалась, однако, значительно менее выраженной, чем можно было ожидать на основании повреждений гипоталамуса. Описано также превращение диких норвежских крыс в ручных после амигдалэктомии [523, 961].
Многочисленные исследования результатов удаления миндалевидного ядра и соседних с ним структур показали важное значение этих частей лимбической системы для формирования эмоций. Тем не менее результаты несколько разочаровывали, так как качественные изменения, возникающие при этих повреждениях, были не одинаковы даже у одного и того же вида. Хотя накопление экспериментальных данных путем исследований на новых видах животных и может помочь нам понять эти сложные вопросы, следует помнить, что функциональное состояние гипоталамуса имеет важнейшее значение для эмоциональной реактивности [90], и поэтому весьма вероятно, что изучение влияния амигдалэктомии и повреждения соседних структур на эмоциональное состояние может дать более четкие результаты у животных, у которых равновесие гипоталамической системы каким-то образом нарушено. Упомянутая выше работа Шрайнера и Клин га указывает на такую возможность.
То особое внимание, которое мы уделяем описанию состояний ярости и превращения животных в ручных, вовсе не означает, что разрушение миндалевидного ядра приводит только к этим" результатам. Грин и др. [400], проанализировав литературу до 1957 г., нашли в ней описание 18 различных симптомов, возникающих при раздражении миндалевидного ядра, и 11 симптомов его разрушения; причем они не претендовали на то, что их обзор является исчерпывающим. Многие из этих симптомов представляют собой результат вегетативных реакций, которые не так просто выявить, и, очевидно, есть основания считать, что синдром Клювера — Быоси и синдром Барда — Маунткастла (т. е. состояния «прирученности» и «ярости») достаточно адекватно характеризуют основные внешние поведенческие изменения, наблвд-
НЕРВНЫЕ И ЭГ-1Д0КРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 113
даемые при повреждениях височной доли и миндалевидного ядра.
При разрушении каудальной области гипоталамуса также происходит превращение диких обезьян в ручных [733]. При этом их лица лишены какого бы то ни было выражения, у них не наблюдается признаков страха или злобы. Раисой и другие исследователи описали при подобном разрушении такие явления, как сонливость, состояние, подобное каталепсии, и отсутствие эмоциональной реактивности [501, 503, 733, 734, 737].
По данным ряда исследователей, животное становится более ручным при повреждении поясной извилины. В первых опытах (в 1888 г.) [116, 486] частичного или полного удаления передней части поясной извилины не было обнаружено заметных изменений эмоционального поведения. Однако в 1939 г. Рансон [734] показал, что удаление поясной извилины у обезьян ведет к понижению эмоциональной реактивности; в более поздних работах это подтвердилось. Исчезновение страха перед человеком, превращение животного в более ручное и безразличие к соседям по клетке были отмечены также после разрушения поля 24 [377, 928, 929]. У людей удаление поясной извилины приводит к снижению враждебности и страха, к поразительному улучшению настроения, появлению чувства удовлетворения и нормализации эмоциональной реактивности [588]; оно оказывает благоприятное влияние в случаях «непреодолимой раздражительности, агрессивности, грубости и возбуждения» [565].
Наряду с изучением влияния соответствующих разрушений гипоталамо-лимбической системы, приводящих к понижению агрессивности, много внимания уделялось опытам по раздражению гипоталамуса, которые описаны 6 гл. IX; в этих опытах было отмечено появление элементов удовлетворения, удовольствия и т. д. Следует отметить также специфическое подавление агрессивности у кошки при раздражении хвостатого ядра [177], безразличие кошки к мыши при гиппокампальных судорогах [30] и указания о возникновении приятных ощущений у больных при раздражении лимбических структур [178, 814].
11-1
ГЛАВА IV
КОРА ГОЛОВНОГО МОЗГА И ГИПОТАЛАМУС
Со времени первых исследований электрической активности мозга, проведенных Бергером, известно, что в коре головного мозга человека и животных возникают ритмические электрические потенциалы, которые можно зарегистрировать как с обнаженной коры, так и через череп. При различных условиях отмечаются различия в напряжении, частоте и форме электрических волн. Электроэнцефалограмма (ЭЭГ) бодрствующего мозга легко отличима от ЭЭГ, зарегистрированной во время сна. Она резко меняется при сенсорных раздражениях, судорогах или при других состояниях.
Раздражение заднего отдела гипоталамуса, вызывающее описанные выше симпатические реакции, одновременно приводит к глубоким изменениям электрической активности коры мозга. Медленные потенциалы покоя в коре сменяются быстрыми колебаниями низкой амплитуды, что обычно считается признаком возбуждения. Далее, если нанести на кору небольшое количество стрихнина, то появляются судорожные потенциалы высокой амплитуды, так называемые стрихнинные пики. На фоне этих пиков очень легко вызвать возбуждение коры путем раздражения гипоталамуса. Стрихнинные пики, которые в состоянии покоя весьма редки и группируются неравномерно, при таком раздражении становятся более частыми и регулярными. Этот эффект наблюдается в любой части коры при раздражении симпатического отдела гипоталамуса (фиг. 12).
Если вместо прямого раздражения гипоталамуса раздражать некоторые рецепторы, то можно обнаружить наряду с возбуждением гипоталамуса диффузную'активацию коры. Так, если ритмически сгибать и разгибать, конечность наркотизированного животного, механически раздражать кожу или применять другие болевые раздражители, возникает реакция пробуждения в гипоталамусе и в коре [59]. Если раздражитель так слаб или наркоз так глубок, что возбуждение не проявляется в одной из структур, оно не наблюдается и в другой. Следует подчеркнуть, что речь идет о генерализованной активации коры, и только это диффузное возбуждение
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ Цб
коры тесно связано с возбуждением гипоталамуса. Очаговое раздражение периферических рецепторов может вызывать локальные изменения ЭЭГ без возбуждения щпоталамуса. Специфические афферентные пути, связывающие каждую рецеиторную область тела с соответствующей ограниченной областью коры, способны к проведению возбуждения даже в тех случаях, когда глубокий наркоз или низкая интенсивность раздражения препятствуют действию диффузной активирующей системы,
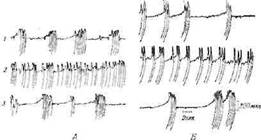
Фиг. 12. Влияние раздражения заднего отдела гипоталамуса на возбуждение коры под действием стрихнина [671].
Электрокортнкограммы. А. Ипсилатеральная задняя сигмовидная извилина.
Б. Ипсилатеральная затылочная доля, I — контроль; 2 — после раздражения
заднего отдела гипоталамуса; .? — контроль спустя 3 мин.
компонентом которой служит каудальный отдел гипоталамуса.
Из этих данных видно, что прямое или рефлекторное возбуждение гипоталамуса вызывает диффузную активацию коры. Кроме того, существует параллелизм между степенью возбуждения гипоталамуса и интенсивностью и длительностью возбуждения коры. Если уровень возбуждения гипоталамуса возрастает, повышается и активность коры; если возбудимость гипоталамуса снижается, как при повреждении заднего гипоталамуса, возбуждение коры исчезает [304, 309, 549].
Ilfi
ГЛАВА IV
Если мы используем зш данные для анализа эмоций, которые, как было показано, связаны с возбуждением гипоталамуса, мы придем к выводу, что эмоциональное возбуждение заключается не только в нисходящем вегетативном разряде гипоталамуса, ведущем к сдвигу в состоянии внутренних органов, но также и в восходящем разряде, вызывающем генерализованное возбуждение коры.
СИМПАТО-АДРЕНАЛОВЫЕ РЕАКЦИИ ПРИ ЭМОЦИЯХ
Классические исследования Кэннона показали, что эмоциональное возбуждение у животных (например, при встрече кошки с лаюшей собакой) сопровождается мощным разрядом всей симпатической системы. Таким образом, все те явления, которые были описаны ранее как следствие прямого или рефлекторного раздражения симпатических центров продолговатого мозга и гипоталамуса, наблюдаются в условиях нормального эмоционального возбуждения у животных. При этих условиях у кошек обнаруживается повышение частоты сердечных сокращений, максимальное расширение зрачка, сокращение третьего века и пилоэрекция. Перистальтика желудочно-кишечного тракта ослабляется, потоотделение усиливается, возрастает число эритроцитов в циркулирующей крови за счет поступления их из селезенки, где они хранятся до момента сокращения гладких мышц под влиянием симпатического импульса.
Мы отметили ранее, что сильное раздражение симпатических центров приводит не только к симпатическому нервному разряду, но также к гиперфункции мозгового вещества надпочечников, о чем свидетельствует сокращение денервированного третьего века и повышение содержания сахара в крови. Целесообразность подобных реакций очевидна. Сокращение третьего века позволяет животному лучше видеть, а следовательно, быстрее реагировать на любое изменение внешней среды. Содержание сахара в крови повышается в результате усиленного расщепления гликогена в печени под влиянием гормона надпочечников. При этом депонированный в печени нерастворимый углевод превращается в растворимую нир-
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЯ 117
кулирующую в крови глюкозу, которая обеспечивает энергию, необходимую для усиленной деятельности скелетных .мышц в условиях нападения и бегства.
Убедительным доказательством того, что именно мозговое вещество надпочечников выделяет такой химический стимулятор, могут служить несколько установленных экспериментально фактов. Один из этих фактов — отсутствие описанных явлений у адреналэктоми-рованных животных. Перерезка симпатических нервов, иннервирующих мозговое вещество надпочечников, или химическая блокада передачи их импульсов также снимает такие реакции, как повышение содержания сахара в крови и сокращение третьего века. На основании изложенного можно сделать вывод, что интенсивное раздражение симпатической системы, искусственное или эмоциональное, активирует как нервные, так и гормональные ракции, и поэтому точнее его следует называть «симпато-адреналовой» или, еще точнее, «симпато-адреномедуллярной» активацией. (Термин «адреноме-дуллярный» поясняет, что кора надпочечников, выделя- о ющая другие гормоны, не участвует в этом процессе ( [133, 296, 306].)
УЧАСТИЕ ПАРАСИМПАТИЧЕСКОЙ СИСТЕМЫ В ЭМОЦИОНАЛЬНЫХ РЕАКЦИЯХ
Многие интересовались вопросом о том, ограничивается ли активация вегетативной системы при эмоциях ее симпатическим отделом или захватывает также и па- j расимпатический отдел. У экспериментальных животных (чаще всего у грызунов) эмоциональное возбуждение нередко сопровождается мочеиспусканием и дефекацией. Это свидетельствует об участии парасимпатической системы в нисходящем вегетативном разряде. Кроме того, активация парасимпатической системы при 'эмоциональном возбуждении проявляется иногда расшире- ' \У нием кровеносных сосудов. Об активации этого отдела вегетативной системы говорят и другие наблюдения. Например, старт-рефлексы у обезьян и новорожденных детей сопровождаются эрекцией, которая связана с резким притоком крови к половому члену. У кроликов эмо-
глава iv
ционалыюе возбуждение сопровождается расширением кровеносных сосудов кожи, усиленной теплоотдачей и. вследствие этого снижением температуры тела.
Эти данные со всей отчетливостью показывают, чго при эмоциях происходит возбуждение парасимнатиче-| ской системы. Тот факт, что у кошки и собаки преобладают симпатические эффекты, а у кролика — парасимпатические, объясняется видовыми различиями в реактивности вегетативной системы. Изучение характера старт-рефлексов показало, что тип эмоции оказывает влияние на соотношение симпатических и парасимпатических компонентов в нисходящем вегетативном разряде- Этот вывод подтверждается наблюдениями над так называемой психической фазой сокоотделения. В работах Павлова по условным рефлексам на собаках было показано, что даже вид пищи может вызвать секрецию слюны и желудочного сока, а также усиление перистальтики желудка и желчного пузыря. Наблюдения показа-\ ли, что если при гневе и страхе преобладают симпатические разряды, то приятные эмоции сопровождаются доминированием парасимпатической активности, ведущей к повышению секреторной и моторной функции желудочно-кишечного тракта [306].
КОМБИНИРОВАННЫЕ СИМПАТИЧЕСКИЕ И ПАРАСИМПАТИЧЕСКИЕ РЕАКЦИИ
Прежде чем подойти к анализу механизмов, определяющих тип вегетативных реакций, наблюдаемых при эмоциях, необходимо рассмотреть еще ряд фактов. Следует иметь в виду, что большинство внутренних органов снабжено как парасимпатическими, так и симпатическими нервами, которые под влиянием электрического раздражения или естественной активации влияют на данный орган в противоположных направлениях. Парасимпатические влияния 'понижают кровяное давление и частоту сердечных сокращений, а симпатические влияния оказывают противоположное действие, однако в некоторых условиях оба отдела вегетативной нервной системы могут активироваться одновременно. Функция органа в таких случаях зависит от относительной эффективности
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 119
возбуждения этих двух отделов. Приведем пример: повышение кровяного давления и частоты сердечных сокращений при эмоциональном возбуждении может быть следствием алгебраической суммации мощного симпатического прессорного и слабого парасимпатического де-прессорного эффекта. С помощью хирургического вмешательства или другими методами можно исключить влияние одного из отделов вегетативной системы и таким образом провести физиологический анализ, показав роль подобного взаимодействия, в связанном с эмоциями изменении сердечного ритма. Так, Шеррингтон наблюдал, что у нормальной собаки шум вызывает учащение сердечного ритма, а после перерезки спинного мозга в верхнем отделе тот же шум приводит к замедлению этого ритма. При такой перерезке пересекаются нервные пути от мозга к ускоряющим симпатическим нервам сердца, но сохраняются связи мозга с парасимпатическим блуждающим нервом. Из этого простого эксперимента можно сделать вывод о том, что эмоциональное возбуждение включает симпатические и пара-; симпатические разряды, причем в норме в организме: преобладают симпатические эффекты.
Этот важный вывод подтверждает ряд других экс иериментов, в которых в качестве индикаторов состояния вегетативной системы использовались химические вещества (гормоны), выделяемые железами внутренней секреции. Выше мы упоминали о том, что мозговое вещество надпочечника выделяет гормоны (адреналин или норадреналин) под влиянием импульсов симпатических нервных волокон, иннервирующих эту железу. Известно также, что раздражение блуждающего нерва (относящегося к парасимпатической системе) стимулирует секрецию инсулина поджелудочной железой. Хотя секреция инсулина в основном регулируется концентрацией сахара в крови, для рассматриваемой нами проблемы важно подчеркнуть, что парасимпатическая система также влияет на секреторную активность островков Лангерганса. Как ни велико значение этого факта, установленное в эксперименте стимулирующее влияние электрического раздражения блуждающего нерва на секрецию инсулина еще не доказывает участия этого механизма при
ГЛАВА IV
физиологических условиях. Однако в приводимых ниже экспериментах были получены такие доказательства.
Для понимания этой работы следует учитывать, что избыточная секреция гормонов мозгового вещества надпочечников приводит к повышению содержания сахара в крови, а избыточная секреция инсулина — к его понижению. При прочих равных условиях сдвиг в содержании сахара является результатом симпати'-4 ческих или парасимпатических разрядов, которые влияют соответственно на мозговое вещество надпочечников или поджелудочную железу. Таким образом, уровень сахара крови может служить индикатором активности симпатической и парасимпатической системы и может быть использован в
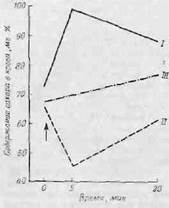
экспериментах, аналогичных опытам Шерриигтона по изучению ритма сердца. Под влиянием внезапного интенсивного звукового раздражения (громкий шум) уровень сахара крови у крыс повышается. У адреналэктомированных крыс тот же раздражитель вызывает гипогликемию. На этом основании можно сделать вывод, что у интактной крысы звуковое раздражение включает одновременно два механизма—один из них вызывает гипергликемию, а другой приводит к гипогликемии. Первый, по-видимому, преобладает, и в конечном счете уровень сахара оказывается повышенным. Поскольку у адоеналэктомнро-
Фиг. 13. Влияние шума на уровень
сахара крови у крысы [306].
/ — в норме; II — после адреналэктомии:
/// — после адреналэктомии и ваготомии.
Стрелкой обозначен момент воздействия
раздражителя.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 121
ванных крыс гипергликемия не развивается, ясно, что она связана с избыточной секрецией адреномедуллярных гормонов в ответ на возбуждение симпатической системы. Гипогликемия, вызываемая тем же раздражителем у адреналэктомироваппых крыс, является результатом секреции инсулина поджелудочной железой под влиянием импульсов парасимпатического нерва. После перерезки блуждающего нерва это влияние исчезает. Если устранить симпатические и парасимпатические влияния, т. е. удалить надпочечники и перерезать блуждающий нерв, то звуковое раздражение не вызовет никаких изменений содержания сахара в крови (фиг. 13). Если вызвать у животного эмоциональное возбуждение, привязав его к доске так, что оно начинает сопротивляться, можно получить такой же результат, что и при действии громкого шума. При встрече с лающей собакой, уровень сахара крови у кошки резко повышается, но если устранить влияние симпатических разрядов на мозговое вещество надпочечников, эффект оказывается противоположным [331]. И в этом случае после перерезки (парасимпатических) блуждающих нервов гипогликемии не наблюдается.
Из этих экспериментов следует, что эмоциональное возбуждение такого типа, которое у человека мы обозначаем как страх, гнев и ярость, вызывает у экспериментальных животных симпатические и парасимпатические разряды; причем в нормальном организме симпатические эффекты преобладают.
АКТИВНОСТЬ СИМПАТИЧЕСКОЙ СИСТЕМЫ ПРИ ЭМОЦИЯХ У ЧЕЛОВЕКА
Широко известно, что при эмоциях происходят выраженные вегетативные сдвиги. Они наиболее очевидны в системе кровообращения — при эмоциональном возбуждении значительно повышается частота сердечных сокращений и кровяное давление. К признакам эмоций относятся также потение ладоней и сухость во рту. Последний симптом представляет собой результат снижения выделения слюны и повышения ее вязкости; подобный эффект можно получить в эксперименте на жи-
ГЛАВА IV
вотных путем раздражения симпатических нервов, подходящих к слюнным железам. Страх сопровождается ,'и другими проявлениями возбуждения симпатической си-/ / стемы, такими, как расширение зрачков, бледность кожи липа вследствие сужения кровеносных сосудов. Состояние тревоги сопровождается снижением активности желудочно-кишечного тракта. Специальные исследования, в частности на больных с фистулами, показали, что при таком состоянии перистальтика кишечника ослаблена. Моторика желудка также ослаблена, и пища задержи-, вается в желудке дольше обычного. Поскольку симпатические нервы оказывают на желудочно-кишечный тракт тормозное влияние, то эти симптомы можно считать показателем усиления симпатической активности. Кроме того, повышается секреторная активность надпочечников. Установлено, что во время экзаменов уровень сахара крови у студентов всегда повышен. В свете ранее описанных экспериментов на животных это свидетельствует об избыточной секреции адреналина мозговым веществом надпочечников, что в свою очередь обусловлено возбуждающим влиянием симпатических нервов. При эмоциях у человека нередко наблюдаются и симптомы повышенной активности парасимпатической системы. При тяжелых формах эмоциональных расстройств "учащаются мочеиспускание и дефекация, а иногда утрачивается произвольный контроль над этими функциями. Слезы — признак гиперфункции слезных желез в результате возбуждающего влияния парасимпатических нервов. Подобная реакция невозможна в случаях повреждения лицевого нерва, несущего парасимпатические импульсы к этим железам. Внезапный шум может вызвать такие парасимпатические реакции, как эрекция и непроизвольная дефекация, наблюдаемые также у обезьян.
Отношения между характером эмоции и природой вегетативных реакций
. 1 Отмечено, что характер эмоции влияет на характер вегетативной реакции у человека. Всем известно, что ог страха человек обычно бледнеет (симпатический эф-
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 123
фект), а от радости — краснеет (парасимпатический эффект). Приятные эмоции, связанные с видом и запахом пищи, ведут к усилению слюноотделения и секреции желудочного сока под влиянием парасимпатических импульсов, тогда как ст,р_ах, действуя через симпатическую систему, вызывает противоположный эффект.
В последние годы наши знания в этой области значительно пополнились благодаря работам Вольфа и сотр. [484, 957], которые изучали реакции желудочно-кишечного тракта и слизистой носа при различных эмоциональных состояниях. Прямое наблюдение за состоянием слизистой желудка на больных с фистулой позволило количественно оценить степень его двигательной и секреторной активности, а также интенсивность кровотока. Эти наблюдения, проводившиеся при различных эмоциональных состояниях, показали, Что повышение секреторной активности и усиление кровотока в этих областях наступает не только под влиянием приятных мыслей о еде, но и при таких эмоциях, как возмущение, негодование и ярость. Было обнаружено, что признаки возбуждения парасимпатической системы не ограничиваются упомянутыми изменениями желудочно-кишечного тракта и слизистой носа; наблюдается, кроме того, гиперемия лица и некоторые проявления со стороны мочевого пузыря и влагалища. Тот факт, что эти изменения не происходят после перерезки (парасимпатического) блуждающего нерва, доказывает, что в основе таких функциональных сдвигов, наблюдаемых при агрессивных состояниях, лежит возбуждение парасимпатической системы. Под влиянием страха происходят изменения иного характера, указывающие на активацию симпатической системы: слизистые бледнеют (сужение кровеносных сосудов), перистальтика ослабляется, секреторная активность снижается.
Хотя эти исследования отчетливо показывают, что различные эмоциональные состояния связаны с разными типами вегетативных реакций, следует остерегаться упрощенного подхода. Негодование и возмущение, как правило, сопровождаются повышением кровяного давления, частоты сердечных сокращений и снижением почечного кровотока. Следовательно, в сосудистой системе
ГЛАВА IV
преобладают симпатические влияния, тогда как в желудочно-кишечном тракте — парасимпатические. Таким образом, для некоторых эмоциональных состояний характерны сложные сочетания вегетативных реакций, а не какая-то' одна из них. Правда, при^^страхе повышенная активность симпатической системы проявляется как циркуляторными симптомами, так и симптомами со стороны желудочно-кишечного тракта, а состояние депрессии характеризуется преобладанием парасимпатического тонуса.
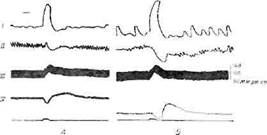
Фиг. 14. Сложный ответ на раздражение заднего отдела гипоталамуса [318].
Характеристика раздражения: 2 в, 99 имп/сек, 0.8 мсек, 10 сек. А. До ва-
готомии. Б. После ваготомии. / — мочевой пузырь; // — двенадцатиперстная
кишка; /// — кровяное давление; IV — денервированное третье веко.
Интересно, что как сложные, так и однородные виды вегетативных разрядов могут быть вызваны в экспериментах на животных путем раздражения гипоталамуса. На фиг. 14 приведен пример разрядов комплексного типа при раздражении гипоталамуса: активность мочевого пузыря возрастает вследствие возбуждения парасимпатической системы, повышение кровяного давления, ослабление перистальтики кишечника и сокращение денервированного третьего века указывают на усиление симпатических разрядов и повышение секреторной активности мозгового вещества надпочечников [318],
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 125
Единый тип реакции на раздражение заднего гипоталамуса в эксперименте показан на фиг. 3 Повышенная симпато-адреналовая активность документируется изменениями со стороны кишечника, мочевого пузыря и сосудов (расслабление мускулатуры кишечника и мочевого пузыря, повышение кровяного давления). Мы указывали, что раздражение переднего гипоталамуса также дает единую реакцию, но в этом случае она носит парасимпатический характер, выражаясь в усилении перистальтики кишечника и снижении кровяного давления. Таким образом, типы вегетативной активности, наблюдаемые при эмоциональных состояниях у человека, подобны тем, которые можно вызвать раздражением гипоталамуса у экспериментальных животных.
Все эти исследования ясно показывают, что различ-f s ные формы эмоций связаны с различными типами вегё-i \у тативных разрядов. Однако это утверждение следует рассматривать в свете других экспериментальных данных. Несомненно, что эмоциональная реактивность различных людей варьирует в широких пределах. Эта вариабельность не ограничивается психологической стороной эмоций, а распространяется и на физиологические проявления. Наблюдения, проводившиеся в бомбоубежищах Англии во время последней войны, показали, что при взрыве бомбы одни бледнели, а другие — краснели. В физиологических терминах это означает, что у одних страх вызывает симпатическую реакцию (сосудов лица), а у других — парасимпатическую.
Тщательные клинические наблюдения дают дальнейшие доказательства индивидуальных различий в характере вегетативных разрядов во время эмоциональных состояний. Ранее мы упоминали, что у большинства здо- ' ровых людей при эмоциональном возбуждении (страх, ярость, напряжение во время экзаменов) происходит по- ] \ вышемие уровня сахара крови, возникающее в резуль- ' тате симпато-адреналовых разрядов. У психически больных такая реакция обычно отсутствует, а при неврозах эмоциональное возбуждение иногда сопровождается гипогликемией [722а]. В свете описанных выше физиологических экспериментов эти данные позволяют считать, что в лорме у людей преобладает симпато-адреналовая
ГЛАЗА !V
система, а при неврозах более реактивна парасимпато-ипсулиновая система. Доказательством правильности такого объяснения может служить факт отсутствия «эмоциональной» гипогликемии после введения,атропина, который блокирует функцию блуждающего нерва. Здесь уместно напомнить, что возбуждение блуждающего нерва при эмоциональных состояниях ведет к повышенной секреции инсулина, который снижает уровень сахара крови.
В результате различных наблюдений, рассмотренных выше, мы приходим к весьма важному выводу о том, что вегетативные разряды, наблюдаемые при эмоциях, определяются двумя факторами: 1) характером возникшего эмоционального возбуждения; 2) степенью уравновешенности симпатических эффектов парасимпатическими.
Определение этой центральной вегетативной реактивности и равновесия систем имеет огромное значение для нашего понимания эмоций в норме и патологии. Эта проблема будет рассмотрена во второй части книги.
Соматические реакции при нисходящих разрядах вегетативных центров
Нисходящие разряды вегетативных центров, в частности гипоталамуса, не ограничиваются вегетативной системой, а воздействуют также и на соматическую. Это хорошо известно из экспериментов на животных, у которых активация вегетативной нервной системы сопровождается агрессивными и оборонительными двигательными реакциями. Даже у таких животных, как кошка, наблюдаются различные двигательные реакции, сопровождающие различные эмоциональные состояния. Так, страх выражается у нее в выгибании спины и мяуканье, а ярость — в шипении, ворчании, попытках укусить и выпускании когтей. Поскольку в обоих случаях вегетативные разряды носят преимущественно симпатический характер, наблюдения позволяют предположить, что соматические реакции при эмоциональных состояниях более многообразны, чем типы вегетативных разрядов.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 127
Это предположение подтверждается изучением эмоций у человека. Мы знаем из повседневного опыта, что выражение лица, зависящее от сокращения мышц в различных комбинациях, по-разному меняется при различных эмоциях. Раздражая электрическим током мышцы лица, Даченн [201] показал, что в выражении радости, горя, страдания и т. д. участвуют различные мышцы. Из этого следует, что сложные комбинации соматической иннервации ответственны за выражение лица при различных эмоциях у человека.
Это, однако, не единственное выражение активности соматической нервной системы, наблюдаемое при эмоциях. Описывая состояние удовлетворения или эмоционального покоя, мы нередко применяем термин «расслабление», а при переходе к состоянию гнева или ярости мы испытываем «напряжение». Этими терминами обозначают определенные физиологические явления. Современные электрофизиологические исследования показывают, что состояние напряжения связано с активацией произвольной (поперечнополосатой) мускулатуры, хотя явных движений при этом может и не быть. В состоянии эмоционального покоя эта активность минимальна или вовсе отсутствует. Дыхательные мышцы также относятся к поперечнополосатым, регулируемым соматической нервной системой. Их активность сразу же меняется под влиянием самых незначительных сдвигов в эмоциональном состоянии и выражается в изменениях ритма и амплитуды дыхательных движений. Вздох, одышка, зевок — общеизвестные примеры таких изменений. Актеры широко пользуются возможностью произвольной регуляции дыхательных движений для передачи эмоций на сцене.
Из экспериментов на животных мы знаем также, что различные типы двигательных поведенческих реакций, выражающих агрессивность, страх, а также приятные переживания (последние менее изучены в этом отношении), имеют подкорковое происхождение; они проявляются даже после удаления больших полушарий. Клинические данные в сочетании с патологоанатомическими указывают на гипоталамус как на основную структуру, Участвующую в организации соматических движений,
ГЛАВА IV
выражающих эмоции. Такое объяснение согласуется с результатами физиологических экспериментов на животных. Кроме того, воздействием на эту область при операциях, производимых под местной анестезией, можно вызвать резкие эмоциональные взрывы, напоминающие психические расстройства.
Выше мы отмечали, что раздражение гипоталамуса вызывает вегетативные и соматические реакции, причем последние выражаются в форме движений, обычно сопровождающих состояния эмоционального возбуждения, такие, как ярость. Здесь мы хотим добавить, что через несколько недель после удаления больших полушарий у кошек можно вызвать сокращение скелетной мускулатуры раздражением гипоталамуса. Поскольку при этих условиях длинные аксоны кортико-бульбарных и кортико-спинальных нейронов подвергаются дегенерации, можно предположить, что движения, вызванные раздражением гипоталамуса, обусловлены возбуждением подкорковых клеток. Это не означает, однако, что данная подкорковая система и в норме не зависит от нервных клеток моторной коры. Напротив, импульсы, возникающие в коре и вызываемые в гипоталамусе, взаимодействуют на уровне продолговатого мозга. Это приводит к мощному усилению возбуждения клеток передних рогов спинного мозга, а следовательно, и к более сильным движениям, чем те, которые возникают при изолированной активации одной из этих систем [671]. Кроме того, как мы видели выше, возбуждение ведет к восходящему разряду, посредством которого активируется вся кора, в том числе и ее двигательная область. Активация моторной коры, вызываемая раздражением гипоталамуса [310] или лимбической системы [732], усиливает разряды пирамидных клеток и, таким образом, приводит к дополнительному усилению движений. Этим объясняется тот факт, что даже при полной потере речи при разрушении центра в лобной коре больные под влиянием эмоционального возбуждения иногда произносят очень трудные слова. Этим же частично объясняется резкое возрастание мышечной силы при некоторых эмоциональных состояниях.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ
КОРТИКО-ГИПОТАЛАМИЧЕСКИЕ СВЯЗИ
Отношения между корой головного мозга и гипоталамусом весьма сложны. С ними следует хотя бы кратко ознакомиться, чтобы лучше понять механизмы эмоций у человека. Полушария мозга в целом и лобная кора в частности оказывают тормозное влияние на гипоталамус. Этот вывод основан на наблюдениях над животными, у которых удалены большие полушария и поврежден обонятельный мозг; у таких животных самые слабые раздражители, даже такие, как поглаживание, могут вызвать реакцию, подобную ярости. Аналогичное «высвобождение» эмоциональных реакций наблюдается у человека на начальной стадии наркоза (поскольку при наркозе сначала подавляется функция коры, а затем подкорковых центров). Кроме того, удаление лобных долей, перерезка путей, соединяющих лобную кору с промежуточным мозгом, или повреждение этих путей при опухолях лобной доли у человека приводят к повышению эмоциональной реактивности, выражающейся в патологической наклонности к бессодержательным шуткам, отсутствии контроля над выражением полового влечения и т. д. Таким образом, «высвобождение» гипоталамуса, очевидно, является важным фактором, лежащим в основе повышенной эмоциональной активности.
Кроме того, на гипоталамус влияет активация некоторых областей коры. Если поместить на поверхность коры, например на затылочную долю, получающую зрительные импульсы от сетчатки глаза, небольшой кусочек фильтровальной бумаги, пропитанной раствором стрихнина, то электрические потенциалы высокой амплитуды возникают не только в непосредственной близости от фильтровальной бумаги, но также и в гипоталамусе [672]. Этот простой опыт можно с равным успехом провести на слуховой и других так называемых сенсорных проекционных зонах коры; это своего рода физиологическая модель эмоционального возбуждения, наступающего под влиянием сенсорных впечатлений. У животных доминирующую <роль в связи с пищевым и половым поведением играет обоняние; у человека главную роль играют зрительные и слуховые восприятия, но и
5-517
ГЛАВА IV
у животных вид пищи вызывает выделение слюны. Несмотря на относительно более важное значение филогенетически более поздних областей мозга у человека, «лимбический мозг», обладающий обширными афферентными и эфферентными связями с внутренними органами, играет чрезвычайно важную роль в организации эмоций.
Эти и подобные им опыты доказали возможность активации гипоталамуса через кору и коры — через гипоталамус. Изменения возбудимости гипоталамуса резко меняют влияние коры на гипоталамус, а также действие гипоталамуса на кору [309]. С этой точки зрения легко понять, что активность гипоталамуса, а следовательно, и эмоциональное состояние сильно влияют иа мозг в целом, а значит, и на мышление.
Можно привести интересный пример гипоталамо-кор-тикальных отношений у человека. На электроэнцефалограмме (ЭЭГ) здорового человека в состоянии покоя видны так называемые альфа-волны — правильные колебания с частотой 10 в 1 сек, регистрируемые с различных точек поверхности черепа (фиг. 15, Б). Под влиянием тревоги эти колебания исчезают .{см. фит. 15, А). То же отмечается при реакции пробуждения, умственном напряжении и сенсорном раздражении. Возбуждение, лежащее в основе этого явления, имеет подкорко-
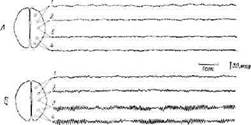
Фиг. 15. Отсутствие альфа-ритма в состоянии тревоги [579].
Д. Состояние тревоги. Б. Норма. Области коры: 1 — лобная; 2 —моторная; 3 — теменная; 4 — затылочная.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 131
вое происхождение; в нем, вероятно, участвует гипоталамус ', как и в описанных ранее экспериментах на животных, в которых было показано, что прямое раздражение или рефлекторная активация гипоталамуса сопровождается диффузным возбуждением коры. Интересно, что иногда у психически здоровых людей при эмоциональном возбуждении возникают волны с частотой 4—7 в 1 сек. Они кажутся подкорковыми по происхождению (гипоталамическими?) и сходными с теми, которые часто регистрируются при патологии (психопатия). Это наблюдение иллюстрирует два важных факта:
1) отсутствие резкой грани между нормой и патологией;
2) глубокое влияние эмоционального возбуждения на работу всего мозга в целом.
ВЛИЯНИЕ ГИПОТАЛАМУСА НА ВАЖНЕЙШУЮ ЖЕЛЕЗУ ВНУТРЕННЕЙ СЕКРЕЦИИ
Гипоталамус и передняя доля гипофиза
Мы уже видели, что раздражение гипоталамуса или эмоциональное возбуждение не только оказывает влияние на вегетативную и соматическую нервную систему, но и активирует железы внутренней секреции — мозговое вещество надпочечников через симпатическую систему и лангергансовы островки поджелудочной железы через парасимпатическую. Однако влияние эмоций на эндокринную систему носит более широкий характер. Многочисленные опыты на животных, а также результаты клинических наблюдений выявили доминирующую роль гипофиза в эндокринной системе; оказалось, что активность большинства желез внутренней секреции, таких, как щитовидная железа, кора надпочечников, мужские и женские половые железы, зависит от гормонов, выделяемых передней долей гипофиза. Эти так называемые «тропные» гормоны специфичны для различных желез внутренней секреции. Если удалить переднюю долю
* * * *
' Аналогичный эффект, был получен с ретикулярной формации ствола мозга (см. гл. IX).
5*
ГЛАВА IV
гипофиза, эти гормоны исчезают и наступает атрофия соответствующих органов, которые сохраняют лишь минимальную долю своей обычной активности.
Для понимания физиологии и патологии эмоций весьма важно подчеркнуть, что перерезка ножки гипофиза, соединяющей его с гипоталамусом, оказывает точно такое же влияние па половые железы и кору надпочечников, как и удаление самой передней доли гипофиза. Поскольку ножка гипофиза почти лишена нервных волокон, полагают, что перенос возбуждающих химических агентов от гипоталамуса к гипофизу осуществляется через кровь. Перерезка ножки гипофиза блокирует этот процесс. Однако кровеносные сосуды этой области быстро регенерируют, так что кровообращение восстанавливается. Исследования показали, что в зависимости от состояния этих кровеносных сосудов раздражение гипоталамуса либо стимулирует выделение тройных гормонов из передней доли гипофиза, либо не оказывает подобного влияния.
В последние годы разработана очень тонкая методика раздражения гипоталамуса. Электроды вводят в определенную область мозга наркотизированного животного с помощью специального стереотаксического прибора. Существуют методы, позволяющие исследователю раздражать гипоталамус у ненаркотизированного животного, находящегося в нормальном состоянии, с помощью дистанционного управления, так что ничто не ограничивает свободного передвижения животного и не влияет на его поведение. Исследования, проведенные с помощью этих методов, показывают, что с образованием различных тропных гормонов связаны определенные области гипоталамуса. Была показана также специфичность повреждения областей гипоталамуса; так, повреждение, блокирующее выработку тиреотропного гормона, не влияет на образование тропных гормонов, и наоборот (фиг. 16).
Рассмотрим далее некоторые последствия^ перерезки ножки гипофиза или специфического повреждения гипоталамуса, влияющего на результат раздражения передней доли гипофиза и секрецию гонадотропных гормонов. У самки кролика овуляция происходит при спаривании,
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 133
так что оплодотворение в матке может произойти позднее. Перерезка ножки гипофиза не влияет на спаривание, но блокирует овуляцию. У грызунов активность яичников носит циклический характер (подобно овари-ально-менструальному циклу у женщин); за ней легко наблюдать по анатомическим изменениям слизистой оболочки влагалища. После соответствующих повреждений

Фиг. 16. Среднесагиттальный разрез через гипоталамус и гипофиз [427а].
Ядра гипоталамуса: / — маммилярные; 2 —заднее; 3 — дорсо-медиалыюе; 4 — вентро-медиальное; 5 — перавентрикулярное; 6 — супраоптическое; 7 — хиазма. Точками обозначены области, раздражение или разрушение которых приводит к изменению соответствующей секреторной активности гипофиза.
гипоталамуса или перерезки ножки гипофиза эти циклические изменения исчезают. У некоторых животных, например у хорька, циклическая половая активность зависит от действия света. При упомянутых выше повреждениях она также нарушается. У некоторых видов животных наблюдается резкое снижение полового влечения при повреждении определенных областей гипоталамуса. Как показывает запись электрической активности гипоталамуса, именно эти области активируются в норме при раздражении наружных половых органов. В свете этих данных можно понять наблюдаемые в клинике случаи нарушения овариальяо-менструального цикла и сложных процессов, лежащих в основе либидо, под влиянием эмоциональных расстройств.
ГЛАВА IV
Недавние исследования указывают на существование в гипоталамусе двойной системы регуляции половых процессов, действующей в противоположных направлениях. Если задний гипоталамус обеспечивает нормальную возбудимость этих процессов, то передний оказывает тормозное влияние. Вот почему повреждение переднего гипоталамуса, при котором снимается тормозное влияние, ведет к преждевременному половому созреванию, а повреждение заднего гипоталамуса задерживает его наступление.
Самые разнообразные раздражения, в том числе боль и условия, ведущие к эмоциональному возбуждению, стимулируют выделение адренотропного гормона. И в этом случае, раздражая гипоталамус, можно вызвать многочисленные проявления действия адренотроп-ных гормонов, а путем повреждения гипоталамуса или перерезки ножки гипофиза можно блокировать эти проявления. Болевые раздражения, эмоциональное возбуждение, повреждение тканей тела, ожоги и т. д. обычно приводят к снижению содержания лимфоцитов и эозипо-филов в крови, однако после соответствующих повреждений гипоталамуса или перерезки ножки гипофиза подобного эффекта не наблюдают. Далее было показано, что при эмоциональном возбуждении концентрация ад-ренокортикотропного гормона (АКТГ) в крови повышается, что стимуляция им коры надпочечников приводит к повышению концентрации вырабатываемого ею гормона в крови и моче. Все это мы рассмотрим более подробно ниже, здесь же ограничимся утверждением, что эмоциональное возбуждение ведет к повышению активности коры надпочечников. В этом отношении существуют выраженные различия между здоровыми людьми и психически больными; это связано, по-видимому, с различной реактивностью гипоталамуса.
Этот краткий анализ данных о секреции адрено- и гонадотропных гормонов свидетельствует о выраженном влиянии гипоталамуса на активность передней доли гипофиза. Однако эмоциональные процессы влияют также на взаимодействие гипоталамуса с задней долей гипофиза.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИЙ 135
Гипоталамус и задняя доля гипофиза
Эти связи отличаются от связей гипоталамуса с передней долей гипофиза. Если передняя доля гипофиза, безусловно, является железой, то задняя состоит из нервной ткани. Нейроны, находящиеся в переднем гипоталамусе, посылают свои длинные аксоны через ножку гипофиза к его задней доле. В телах этих нейронов вырабатывается ряд гормонов, которые переносятся через их волокна к заднему гипофизу, где они накапливаются (см, [42]). Шаррер и Шаррер [799] обнаружили перенос гормонов в своих гистологических исследованиях (фиг. 17). [306, 427, 439].
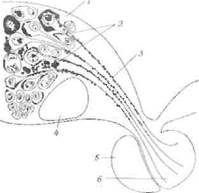
Фиг. 17. Схема, иллюстрирующая образование нейрогормонов в гипоталамусе и их перенос к нейрогипофизу у позвоночного животного [799].
' — преоптичсское ядро; 2 — коллоид; 3 — преоптико-гипофизарный тракт; 4 — хиазма; 5 — передняя доля гипофиза; б —задняя доля гипофиза.
Одним из таких нейросекретов является антидиуретический гормон (АДГ). Физиологически его значение сводится к поддержанию нормального осмотического
ГЛАВА IV
давления в жидких средах тела путем регуляции обратного всасывания воды в канальцах почек. Для нас этот гормон представляет интерес тем, что уровень его секреции может служить показателем активности переднего гипоталамуса при состояниях, не связанных с изменением осмотического давления. Например, его секреция возрастает под действием боли или эмоционального возбуждения. Мирский [651] показал, что болевое раздраже-пые повышает секрецию АДГ. Боль, возникающая при произвольном сокращении ишемизированной мышцы, сопровождается повышением концентрации АДГ в крови. Психологический и физиологический стрес, сопровождающий хирургическое вмешательство, приводит к тому же эффекту. Верни [905а] показал, что ярость, вызванная у собаки внезапным звуковым раздражением, приводит к такому же резкому прекращению мочеотделения, как и введение АДГ. Даже помещение на несколько минут в незнакомую обстановку вызывает у крыс повышение секреции антидиуретического гормона (см. также [654—656]).
Эти данные представляют большой интерес для нашей концепции, касающейся организации эмоций. Мы указывали выше, что передний и задний гипоталамус несут основную ответственность за парасимпатические и симпатические эффекты, возникающие при эмоциональном возбуждении. Наблюдения, описанные в этом разделе, подтверждают и расширяют наш общий вывод, показывая, что передний (парасимпатический) и задний (симпатический) отделы гипоталамуса участвуют также в секреции гормонов передней и задней долей гипофиза.
Известны многие другие факты, касающиеся гипота-ламо-гипофизарных связей вообще и при эмоциональном возбуждении в частности. Ядра переднего гипоталамуса вырабатывают еще один гормон — окситоцин, который стимулирует сокращение матки во время родов. Кроме того, он выделяется во время полового сношения и способствует проникновению сперматозоидов в матку. Лактация также находится под контролем гипоталамуса и гормонов передней и задней долей гипофиза. Из-за недостатка места мы не можем здесь вдавать-
РВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ
Ся в детали. Однако приведенных данных достаточно, чтобы показать, что возбуждение гипоталамуса при общих типах эмоционального возбуждения, таких, как страх, тревога, негодование, а также при особых эмоциональных состояниях, возникающих при половом сношении, приводит к глубоким изменениям активности различных желез внутренней секреции через активацию гипофиза.
ЗАКЛЮЧЕНИЕ
Декортицированное животное часто реагирует яростью на самые «безобидные» раздражители. Такие реакции плохо направлены, прекращаются сразу же по прекращении действия раздражителя и, по-видимому, не сопровождаются эмоциональным ощущением. Низкий порог этих реакций после декортикации позволяет говорить о «высвобождении» низших центров из-под тормозного действия коры. Дополнительным свидетельством в пользу такого механизма служит высвобождение «эмоциональных» реакций на начальных стадиях наркоза у человека и повышение эмоциональной реактивности после повреждения лобных долей.
Раздражение заднего гипоталамуса вызывает такие же реакции, и если оно производится у ненаркотизиро-ванного, интактного животного, то может вызвать отчетливую реакцию ярости с хорошо направленным агрессивным или защитным поведением. Таким образом гипоталамус, по-видимому, и олицетворяет те «низшие центры», о которых мы упоминали. Тот факт, что выражения эмоций у ненаркотизированных животных хорошо направлены, а также то, что повышенная возбудимость сохраняется и после прекращения раздражения, показывает, что данная эмоция не только выражается, но и ощущается.
Раздражение рецепторов вызывает возбуждение специфических проекционных зон коры без параллельного возбуждения гипоталамуса и без признаков эмоций, особенно при локализованном и слабом раздражении. В состоянии глубокого наркоза даже сильные периферические раздражения возбуждают только специфические
ГЛАВА IV
проекционные пути — диффузные афферентные системы гипоталамуса и ретикулярной формации не реагируют, и никаких признаков ощущения или выражения эмоций не отмечается. Однако у ненаркотизированного животного или при поверхностном наркозе тот же раздражитель вызывает возбуждение заднего гипоталамуса, что приводит к диффузному возбуждению коры. При условиях, повышающих или понижающих возбудимость гипоталамуса, диффузная реакция коры также соответственно усиливается или ослабляется, а это весьма важный фактор в эмоциональной реактивности.
Между лимбическим мозгом и гипоталамусом существует тесная связь. У животных различных видов эмоциональное состояние может меняться в широких пределах при раздражении или повреждении миндалевидного ядра, поясной извилины и гинпокампа.
Повышенная эмоциональность с проявлениями злобы и агрессивности в поведении, возникающая при разрушении или раздражении элементов гипоталамо-лимби-ческой системы, а также сниженная эмоциональность, спокойствие и усиление реакций, выражающих удовольствие, при разрушениях и раздражении других элементов той же системы заставляют считать, что именно здесь локализуются структуры, имеющие первостепенное значение для механизмов эмоций. Было бы неразумно считать, что эмоции зависят только от этой системы или что эта система участвует только в организации эмоций.
Однако в свете имеющихся данных было бы столь же неразумно недооценивать роль гипоталамо-лимбиче-ской системы в эмоциональных процессах. Когда мы говорим о гипоталамо-кортикальных разрядах, мы имеем в виду диффузную активацию новой коры, а через нее — лимбической системы, и наоборот.
В эмоциональных реакциях участвуют как симпатический, так и парасимпатический отделы вегетативной нервной системы, посредством своих восходящих и нисходящих разрядов они вносят свой вклад в ощущение и выражение эмоций. Их влияния не ограничиваются нервными воздействиями, но касаются и сдвигов эндокринного равновесия.
НЕРВНЫЕ И ЭНДОКРИННЫЕ КОРРЕЛЯТЫ ЭМОЦИИ 139
В интактном организме при эмоциональных состояниях обычно доминируют симпатические влияния, но имеются данные, показывающие, что разным видам эмоций соответствуют различные типы вегетативных разрядов и при некоторых эмоциональных состояниях доминирует парасимпатическая система. В других случаях вегетативные реакции носят более сложный характер, так что в функциональной активности одних органов преобладает симпатическая система, а других — парасимпатическая. Эти различные по характеру реакции, наблюдаемые при различных эмоциональных состояниях у человека, можно вызвать и в эксперименте путем раздражения гипоталамуса. Такое раздражение иногда вызывает «единый» тип вегетативной реакции — симпатическую или парасимпатическую; в других случаях реакция бывает «комплексной», так что, если использовать в качестве индикатора один орган, можно ошибочно обозначить ее как симпатическую или парасимпатическую в зависимости от того, какой орган избран, тогда как на самом деле она является смешанной. Индивидуальные различия не позволяют точно предсказать природу вегетативной реакции на данную ситуацию, вызывающую эмоциональную реакцию. Даже у одного и того же индивидуума эмоциональная и вегетативная реакции на постоянный раздражитель меняются со временем и под влиянием изменений внутреннего состояния. Тем не менее можно прийти к правомерному выводу, что форма вегетативного разряда, сопровождающего эмоцию, определяется ее типом, хотя определенную роль играет и предшествующее ей состояние вегетативного равновесия.
Степень активности различных симпатических и парасимпатических центров, очевидно, влияет на характер ощущений и выражения эмоций. Последние определяются активностью скелетных мышц, а эти соматические эффекты, как и висцеральные, модулируются нисходящими разрядами гипоталамуса. Сокращения соматических мышц могут быть вызваны раздражением гипоталамуса даже после двухстороннего удаления полушарий и дегенерации кортико-бульбарных и кортико-спиналь-ных нейронов. Если же кора и ее нисходящие проекционные пути не повреждены, раздражение гипоталамуса
ГЛАВА IV
модулирует соматическую активность через восходящие гипоталамо-кортикальные разряды. Такое влияние имеет место при эмоциональном возбуждении; возникающим при этом облегчением в коре можно объяснить, например, описанное .выше парадоксальное явление временного восстановления речи в случаях ее стойкой потери.
Кроме активации коры со стороны гипоталамуса и тормозного влияния коры на гипоталамус, между этими двумя структурами существует другая форма взаимодействия. Возбуждение некоторых областей коры приводит к активации гипоталамуса. Благодаря таким двухсторонним связям кора и гипоталамус способны возбуждать друг друга. Несомненно, это имеет важное значение для реакции пробуждения и усиления эмоциональных процессов.
Внутренние состояния, которые в конечном счете определяют все наши ощущения и поступки, находятся под влиянием не только нервной, но и эндокринной системы. Гипоталамус обеспечивает тесную связь между двумя системами. Он не только регулирует функцию симпатического и парасимпатического отделов вегетативной системы, а через них функцию эндокринных желез, но и определяет секреторную активность «главной» эндокринной железы — гипофиза. Гормоны вырабатываются в самом гипоталамусе, что в значительной степени определяет секреторную активность передней и задней долей гипофиза, глубоко влияя, таким образом, на эндокринное равновесие всего организма в целом.
Короче говоря, как нервные, так и эндокринные .факторы, участвующие в эмоциях, интимно связаны с активностью гипоталамуса, а через него с лимбической системой, корой головного мозга, эндокринной системой и даже скелетной мускулатурой.
ЧАСТЬ ВТОРАЯ
ОБОСНОВАНИЕ И ПРИМЕНЕНИЕ МЕТОДОВ ИССЛЕДОВАНИЯ ГИПОТАЛАМУСА
ГЛАВА *
ФАКТОРЫ,ОПРЕДЕЛЯЮЩИЕ АКТИВНОСТЬ
СИ МП А ТИЧЕСКИХ
И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ
В предыдущем разделе было показано, что эмоциям всегда сопутствуют изменения активности вегетативной нервной системы. Мы разделяем с другими авторами убеждение в том, что для понимания физиологии и патологии эмоций необходимо точное знание характера этих изменений. Однако мы причисляем себя к противникам теории Джеймса — Ланте, согласно которой вегетативные сдвиги являются причиной эмоций. Вряд ли сейчас кто-нибудь серьезно станет утверждать, что мы печальны, потому что плачем. Действительно, не периферический процесс, локализованный в слезных железах, а центральный процесс, интимно связанный с состоянием гипоталамуса, является первичной причиной, лежащей в основе эмоциональной реакции. Он находит свое выражение в характерных вегетативных и соматических разрядах, которые вместе с кортикальными реакциями составляют эмоцию.
Эти вегетативные сдвиги тесно переплетаются с активностью коры мозга. Гипоталамус может вызвать сдвиги вегетативной активности в ответ на нервные импульсы, которые он получает от коры. Кора в свою очередь получает импульсы от гипоталамуса. Если принять, что личность определяется активностью мозга в целом (в том числе и типом эндокринного равновесия, который в свою очередь определяется активностью мозга), не удивительно, что эти реакции обнаруживают зависимость от поведения и личности. Так, многие авторы отмечали, что интенсивность вегетативных разрядов возрастает при фрустрации. В таких случаях, как правило,
ГЛАВА V
появляется сердцебиение, потеют ладони, повышается кровяное давление. Если же испытуемый или больной имеет возможность выразить в .поведении реакцию, соответствующую ситуации, вегетативные реакции минимальны [170]. Лейси и Лейси [559] на основании тщателыных и разносторонних исследований высказали предположение, что характерные черты личности, например импульсивность, связаны с типом вегетативной реактивности, проявляющейся при отдыхе и работе. Так, у «импульсивных» личностей даже в покое отмечаются ритмические колебания интенсивности потоотделения на ладонях и частоты сердечных сокращений. У других лиц в тех же условиях подобных колебаний не наблюдается. Эти и аналогичные исследования показывают, что интенсивность и частота нервных разрядов, возникающих в вегетативных центрах .мозга, особенно в центрах гипоталамуса, отражают количественные различия в реактивности у отдельных людей или у одного и того же человека в разных условиях. В подтверждение этого предположения можно привести тот факт, что вещества, которые .непосредственно повышают реактивность вегетативных центров, такие, например, жак метразол, непосредственно и рефлекторно вызывают ритмические вегетативные реакции. Так, третье веко кошки, которое обычно во время наркоза расслаблено, начинает ритмически сокращаться после инъекции метразола [333]. Особенно интересно подчеркнуть, что подобный эффект и аналогичные ему колебания кровяного давления возникают после инъекции минимальных количеств другого возбуждающего вещества, стрихнина, в симпатический отдел гипоталамуса. Таким образом, вегетативная активность, очевидно, отражает состояние вегетативных центров, в частности гипоталамуса.
Это объяснение приложимо равным образом и ,к парасимпатической системе, поскольку у разных индивидуумов в состоянии эмоционального возбуждения наблюдаются выраженные количественные различия в ее реактивности. Обморочное состояние вследствие резкого падения кровяного давления, сопровождающегося замедлением сердечного ритма, часто возникает при виде крови или в состоянии страха, когда человеку что-то
АКТИВНОСТЬ СИМПАТИЧЕСКИХ И ПАРАСИМПАТИЧЕСКИХ ЦЕНТРОВ 145
угрожает. Симптомы повышенной реактивности парасимпатической системы часто наблюдаются у невротиков и лип с навязчивыми состояниями [895].
Приведенные выше примеры показывают, что типы эмоциональных вегетативных реакций тесно 'связаны с особенностями личности. Если бы мы научились изменять вегетативную реактивность на уровне гипоталаму-j са, это «южно было 'бы использовать для лечения невро-1 зов и психозов. Прежде чем проводить дальнейший анализ этих проблем, мы на1мерены обсудить исследования, иллюстрирующие возможность изменения реактивности гипоталамуса у экспериментальных животных. Если мы примем, что в основе ощущения и выражения эмоций лежат гипоталамические разряды, то мы тем самым признаем, что открытие путей регуляции реакций гипоталамуса равносильно обнаружению способов регуляции самих эмоций. Потенциальное значение такой регуляции для психического (да и физического) здоровья со всеми его последствиями для личности и для общества трудно переоценить. Мы считаем, что весьма важные первые шаги к пониманию регуляции реактивности гипоталамуса и связанных с нею эмоциональных реакций уже сделаны, и мы обсудим их ниже. Экспериментальные данные, .которые мы опишем, не дают полного ответа на вопрос о путях регуляции гипоталамуса — до этого еще далеко. Мы можем только надеяться, что эти данные окажутся направляющими для дальнейшей исследовательской работы и укажут пути ее плодотворного развития.
ПОНЯТИЕ «НАСТРОЙКИ» В НЕРВНОЙ СИСТЕМЕ
Из повседневного опыта известно, что реакция человека на данную ситуацию в значительной степени зависит от его умственного, физического и эмоционального состояния. Можно сказать, что человек «настроен» реагировать определенным образом. Вегетативные реакции на данный раздражитель могут быть то в основном симпатическими, то в основном парасимпатическими. Можно сказать,, что вегетативная система «настроена» таким образом, что тот или иной ее отдел оказывается макси-
146 /ГЛАВА V
мально чувствительным. Раздражитель, обладающий способностью возбуждать оба отдела, может вызывать реакции, различающиеся^ степени участия в «их .каждого из этих центров. Сенсибилизация вегетативных центров была обозначена термином настройка [310], так что принято говорить о симпатической или парасимпатической настройке. В наш век электроники термин настройка обычно ассоциируется с настройкой приборов на резонансную частоту. Поэтому мы должны подчеркнуть, что в этих рассуждениях этот термин не имеет отношения к частоте и указывает только на сенсибилизацию или облегчение определенных центров головного (и спинного) мозга.
Настройка в соматической нервной системе
Рефлекторные движения, такие, «ак сгибание задней конечности в ответ на укол в область подошвы, как правило, легко воспроизводятся. Качественно и количественно ответ остается постоянным, если то же раздражение наносится с достаточными интервалами. Это можно наблюдать на фоне глубокого наркоза, при котором высшие отделы центральной нервной системы невозбудимы, или на спинальном животном, у которого головной и 'спинной мозг разобщены. Это так называемые спинальные рефлексы, которые свободны от каких бы то ни было влияний со стороны головного мозга, что делает их значительно более устойчивыми и надежными. Благода
Дата добавления: 2014-12-10; просмотров: 1269;