Эффект отдачи и разряд последействия
После прекращения действия раздражителя, тормозящего текущую активность', последняя часто усиливается— это так называемый послетормознои эффект отдачи. Он зависит от разряда последействия, который служит как бы продолжением влияния раздражения после того, как само действие раздражителя прекратилось. Если раздражитель оказывает как возбуждающее, так и тормозное влияние на мотонейроны и если возбуждающий разряд последействия сохраняется в течение более длительного времени, чем тормозной, возникает эффект отдачи.
Возбуждающий и тормозной разряды последействия могут быть весьма продолжительными вследствие существования в центральной нервной системе открытых (множественных) цепей и замкнутых реверберируюишх контуров (фиг. 1). Можно видеть, что при наиболее
* * * *
1 Торможение не имеет смысла, если оно не снижает активность, или возбудимость.
ПРИНЦИПЫ НЕРВНОЙ ИНТЕГРАЦИИ 45
прямых связях между афферентными и эфферентными нейронами имеет место минимальная синаптическая задержка. Импульсы, идущие по менее прямым путям, достигают мотонейронов после продолжительного лаг-периода, причем некоторые приходят к мотонейронам уже после прекращения раздражения и передачи импульсов по более прямым путям. Этот эффект сильнее

Фиг. 1. Два основных вида цепей в центральной нервной системе.
А. Открытая (множественная) цепь. В. Замкнутая цепь (реверберирующий контур). Второй контур на фиг. Б (тонкие линии) иллюстрирует возможность взаимодействия многих цепей и поддержание активности в течение значительного времени.
выражен в реверберирующих контурах, чем в открытых цепях, поскольку для первых характерно наличие обратных связей, обеспечивающих повторное самовозбуждение. С такой реверберацией, происходящей в мозге, связывают кратковременную память. До тех пор пока разряд последействия продолжается, он влияет на возбудимость центральных клеток и, таким образом, участвует в регуляции соотношения между входными и выходными сигналами в центральной нервной системе. Более того', возможно, что по крайней мере ,в мозге изменения возбудимости не обязательно прекращаются, когда кончается разряд последействия, но могут со-
4fi
ГЛАВА I
храниться в какой-то форме неопределенно долгое время. Очевидно, не существует более адекватного объяснения для существования долговременной .памяти, которая не исчезает даже после резкого снижения активности мозга, как, /например, .при глубоком наркозе или коме.
Фракционирование
Ни один афферентный нерв не способен вызвать возбуждение .всех мотонейронов данной мышцы. Различные афферентные .нервы контролируют отдельные части мышц (фракции); именно это и имеют в виду нейрофизиологи, когда говорят о «фракционировании». Некоторые нервы контролируют большие фракции .мышц, другие — значительно меньшие. Кроме того, существует центральное перекрытие, в результате которого каждый мотонейрон возбуждается несколькими афферентными нервами. Это центральное перекрытие зон ведет к окклюзии.
Окклюзия
Окклюзию можно назвать эффектом «занятой линии». Если одно раздражение .вызвало максимальное возбуждение центральных нейронов, дополнительные раздражения ничего не добавляют к возникшей реакции. Например, раздражение любого из двух афферентных нервов может .вызвать значительное рефлекторное сокращение мышцы. Если сокращение, возникающее в результате раздражения обоих нервов, идентично тому, которое можно получить при изолированном раздражении каждого из них, это будет указывать на 100%-ную окклюзию, зависящую от полного перекрытия зон. Активность мотонейронов регулирует тот нерв, который первым их возбуждает. Такая степень перекрытия вряд ли вероятна. Значительно более обычный результат совместного раздражения нервов выражается в возникновении сокращения, которое будет несколько больше, чем .при раздражении каждого из нервов в отдельности, но меньше, чем сумма сокращений, вызы-
ПРИНЦИПЫ НЕРВНОЙ ИНТЕГРАЦИИ
васмых их раздельным раздражением. Степень окклюзии можно оценить, если вычесть из суммы «апряже-ний, вызываемых каждым первом в отдельности, напряжение, возникающее в результате одновременного их раздражения.
Количественная рефлекторная регуляция эфферентных нейронов
Рефлекторная реакция находится в количественной зависимости от интенсивности раздражения и размеров раздражаемого рецепторного поля. В определенных пределах чем сильнее раздражение, тем сильнее реакция.
Интенсивность раздражения, действующего на рефлекторный центр, определяется числом афферентных импульсов, приходящих к нему в единицу времени. (В данном случае мы не будем учитывать тормозных влияний.) По мере повышения интенсивности периферического раздражения соответствующим образом возрастает число афферентных импульсов в секунду, что приводит к более сильному раздражению рефлекторного центра. Возникает вопрос, каким образом интенсивность центрального раздражителя определяет интенсивность двигательной реакции.
Подобно тому как при повышении интенсивности периферического раздражения активируется больше рецепторов и отдельные рецепторы разряжаются с большей частотой, более сильное центральное раздражение охватывает больше мотонейронов и повышает частоту их разрядов. Более выраженная суммация возбуждающих влияний в центральной нервной системе при действии более мощного притока афферентных импульсов зависит от накопления необходимых концентраций центрального возбуждающего медиатора в более обширной области или в большем количестве. Мы уже указывали, что процесс возбуждения подчиняется закону «все или ничего»; следовательно, пи импульсы эфферентных нервов, «и импульсы в мышечных волокнах не меняются по амплитуде при изменении интенсивности центрального раздражения. (Напряжение токов действия не зависит от силы раздражения.) Однако степень
ГЛАВА I
суммации в рефлекторных центрах определяет силу реакции эффектора -путем регуляции числа участвующих элементов (вовлечение) и частоты разрядов мотонейронов.
Влияние вовлечения остается в основном одинаковым (Независимо от того, вызывается ли ответ эффектора рефлекторным путем «ли раздражением двигательного нерва. Иначе обстоит дело с частотой разрядов мотонейронои при рефлекторном й прямом раздражении (раздражение двигательного .нерва). В последнем случае волокла двигательного нерва возбуждаются синхронно, тогда как центральная нервная система никогда (если не считать судорожных разрядов) не посылает синхронных импульсов. Пожалуй, наиболее близок к синхронному разряд мотонейронов при коленном рефлексе, однако и в этом случае большинство мотоиейро-нов дает повторные разряды, причем каждый из них в виде коротких залпов, следующих с особой частотой. Таким образом, усиленное напряжение, наблюдающееся при -повышении частоты раздражения двигательного нерва, не отражает .истинной картины влияния частоты на реакцию мышцы в нормальных условиях. В каждый данный момент напряжение является суммой напряжений активных единиц. Даже когда все мышечные волокна отвечают лишь одиночными сокращениями с полным расслаблением между сокращениями, напряжение сухожилия может поддерживаться на вполне постоянном уровне. Это зависит от нескольких факторов: 1) длительность одиночного сокращения значительно превышает длительность вызвавшего его мышечного импульса, так что возможность суммации напряжений множества волокон выше, чем можно предположить на основании определения токов действия; 2) поскольку импульсы, поступающие в различные мышечные единицы, не синхронизированы, одни волокна достигают максимального сокращения, в то время как другие либо еще укорачиваются, либо уже удлиняются; 3) эластичность мышечной ткани также способствует сглаживанию натяжения, передающегося на сухожилие. С увеличением частоты сокращений повышается вероятность суммации. Если некоторые элементы отвечают чаетич-
ПРИНЦИПЫ НЕРВНОЙ ИНТЕГРАЦИИ 49
ным или полным тетанусом, вероятность суммации их сокращений с реакциями других волокон еще более возрастает.
Мы выбрали мышечные реакции для иллюстрации механизма регулирующего влияния центров спинного и головного мозга на величину реакции эффекторов. Все сказанное относится к регуляции функций любых эффекторов и иллюстрирует механизмы, посредством которых осуществляется общеизвестная зависимость между интенсивностью раздражения ,и силой рефлекторной реакции.
Реципрокная иннервация
На принципе реци-прокной иннервации мы остановимся особо, главным образом в связи с отношениями между симпатическими и парасимпатическими центрами вегетативной регуляции. Но общее представление об этом принципе удобнее дать на примере рефлекторной регуляции функции скелетных мышц.
Центры, регулирующие антагонистические функции, соединены между собой вставочными нейронами, так что повышение активности одного из центров сопровождается понижением возбудимости центра, вызывающего противоположный эффект. Например, если раздражение афферентного нерва вызывает рефлекторное сгибание конечности, происходит торможение мотонейронов соответствующих мышц-разгибателей. Предполагается, что все афферентные нервные волокна, входящие в центральную нервную систему, оказывают возбуждающее влияние на нейроны. При реципрокной иннервации приходящие сигналы, вероятно, возбуждают два вида вставочных нейронов; одни из них оканчиваются на мотонейронах сгибателей, где выделяется возбуждающий медиатор, а другие контактируют с мо-гонейронами разгибателей и выделяют тормозной медиатор.
Реципрокная иннервация не ограничивается мышцами одного сустава или даже одной конечности, но распространяется также на контралатеральные конечности, где реакция также носит антагонистический характер.
ГЛАВА I
Так, при широко известном перекрестном разгибатель-ном рефлексе контралатеральное разгибание сопровождается ипсилатеральным сгибанием.
Такой характер взаимодействия между центрами типичен для всей центральной нервной системы. Например, центры вдоха и выдоха в продолговатом мозге также находятся -в реципрокных взаимоотношениях: при возбуждении одного происходит торможение другого. Центры ускорения и замедления сердечной деятельности связаны такими же взаимоотношениями. Число подобных примеров можно умножить, но принцип везде один. Указав ранее, что важнейшей характеристикой интеграции является способность осуществлять в каждый данный момент времени один основной акт, мы должны теперь подчеркнуть, что реципрокная иннервация имеет первостепенное значение для обеспечения этой способности. Сходные механизмы лежат и в основе доминирования определенных рефлексов; так, когда доминируют половые рефлексы, другие рефлекторные акты подавляются.
Хотя принцип реципрокности и имеет, большое значение, не следует думать, что он является основным фактором интеграции. Его можно преодолеть, например, при произвольном напряжении конечности, когда сокращаются как сгибатели, так и разгибатели и суставы оказываются неподвижными, или при прыжке с одновременным сокращением мышц-разгибателей обеих ног.
ЗАКЛЮЧЕНИЕ
Все функции человека и животных связаны с изменениями в структурах организма. Жизнь требует расходования энергии для поддержания организации, без которой живой организм был бы лишен способности поддерживать возбудимость и соответствующим образом приспосабливаться к изменяющимся условиям окружающей среды. Это приспособление осуществляется главным образом эффекторами — мышцами и железами, а их деятельность зависит от периной системы, которая собирает информацию, касающуюся окружаю*
ПРИНЦИПЫ НЕРВНОЙ ИНТЕГРАЦИИ
Si
щей среды, разрабатывает программу действия и направляет к эффекторам сигналы, регулирующие их активность. Различные виды рецепторов, каждый из которых высокочувствителен к определенной форме энергии и направляет сигналы по определенному нервному пути, обеспечивают первичный, анализ изменений в окружающей среде. Вид раздражения и расположение раздражаемой рецепторнои зоны определяются главным образом на основе топографического разделения афферентных путей. Рецептор кодирует различия в интенсивности раздражения различным числом афферентных импульсов в единицу времени. Центральные рефлекторные механизмы на основе получаемой ими количественной и качественной информации определяют число, вид и расположение требующихся эффекторных элементов и интенсивность, с которой они должны реагировать. Реципрокные отношения между антагонистическими рефлекторными центрами исключают возможность хаотического и неэкономного соперничества эффекторов. Вместо этого в результате взаимодействия различных частей нервной и эндокринной системы происходит интеграция функций, позволяющая организму осуществлять одно основное действие в каждый данный момент времени.
ГЛАВА ''
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
МАКРОСТРУКТУРА
Б общем плане строения нервной системы принято различать центральную нервную систему (головной и спинной мозг) и периферические нервы, отходящие от нее ко всем областям тела. Существуют черепномозго-вые « спинномозговые парные нервы.
Как центральная, так и периферическая .нервная система состоит из нервных клеток, или нейронов, которые чрезвычайно разнообразны по размерам и форме. Как правило, нейрон имеет один длинный протоплазма-тический отросток, или нервное волокно, по которому проводятся нервные импульсы. Волокна, которые оканчиваются в пределах центральной нервной системы,— будь то афферентные волокна, идущие с периферии, или отростки вставочных нейронов — многократно ветвятся и образуют множественные синаптические контакты с другими нейронами. Тела мотонейронов, иниер-вирующих скелетную мускулатуру, лежат в центральной нервной системе, от которой к мышце идет длинный аксон, который тоже ветвится и заканчивается нервно-мышечной пластинкой во многих мышечных волокнах. Кроме одиночного длинного волокна, нервная клетка обычно имеет несколько более коротких отростков, называемых дендритами и служащих для увеличения поверхности синаптических контактов.
Тела афферентных, .или сенсорных, нейронов лежат в ганглиях, расположенных вне центральной .нервной системы, но вблизи нее; импульсы от рецептора • (или рецепторов), расположенных дистально от ганглия, передаются клеткам по длинным отросткам. Функцио-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
нально волокна, проводящие импульсы к телу клетки, относят к дендритам, а те, которые проводят импульсы от тела клетки,— к аксонам. Смешанные нервы состоят из многочисленных волокон, и если классифицировать эти волокна строго по функции, то одни из них следует отнести к аксонам, а другие — к дендритам. Однако морфологически они неразличимы, и те и другие способны проводить импульсы в любом направлении, но в норме этого не происходит; объясняется это характером их изоляции от источников раздражения и более низкой чувствительностью самого волокна по сравнению с рецепторами в дистальной части афферентных нервов и дендритам.и и телом эфферентных нейронов. Все волокна периферических нервов независимо от их функции обычно называют аксонами.
СЕГМЕНТАРНЫЙ И НАДСЕГМЕНТАРНЫЙ ОТДЕЛЫ ЦЕНТРАЛЬНОЙ НЕРВНОЙ СИСТЕМЫ
Позвоночное животное состоит фактически из ряда сегментов, каждый из которых имеет свою рефлекторную дугу, осуществляющую функциональную связь между рецепторами и эффекторами. У некоторых беспозвоночных сегментарное строение отчетливо видно даже у взрослых особей. Примером может служить дождевой червь. У человека явные признаки сегментарной организации отсутствуют, однако строение позвоночного столба определенно отвечает принципу сегментарное™. Каждому сегменту спинного мозга соответствует пара нервов, которые связывают нервную систему с отдельными областями тела; связь эта сложилась на ранних этапах эволюции живых существ и сохранилась у высших животных, хотя у последних иннервируемые структуры сильно сместились по отношению к их исходному положению вблизи соответствующего сегмента спинного мозга.
Простейшую форму интегративной деятельности центральной нервной системы представляет рефлекс, и каждый сегмент располагает всеми необходимыми механизмами для его осуществления. К таким механизмам относятся рецепторы, афферентные волокна, идущие к спин-
ГЛАВА It
ному мозгу, эфферентные Волокна к эффекторному органу, сам эффектор и важнейшие синаптические связи между афферентными и эфферентными нейронами в центральной нервной системе. Все эти элементы вместе составляют простую рефлекторную дугу. Как было отмечено выше, такая рефлекторная дуга представляет собой моно-синаптическую, или двухнейронную, организацию, поскольку афферентный нейрон образует синапс непосредственно с эфферентным. В большинстве дуг между афферентным и моторным нейронами имеются вставочные нейроны, так что рефлексы мышц на растяжение в сущности являются единственными, имеющими моноси-наптическую дугу. В сегментарной системе заложены механизмы для таких рефлексов, как коленный, сгибатель-ный, перекрестный разгибательный и многие другие. Большинство сегментарных рефлексов «а самом деле обеспечивается не одним сегментом, а несколькими связанными. Однако реакции осуществляются через наикратчайшие пути и взаимодействие между сегментами не обязательно.
Некоторые из рефлексов, осуществляемых через черепно-мозговые нервы, принципиально сходны со спинномозговыми сегментарными рефлексами, хотя для этих нервов не характерно правильное размещение, типичное для спинномозговых нервов,— слияние сегментов и неравномерный характер роста ствола мозга приводит к утрате явной сегментарности. Примером такого рефлекса может служить зрачковый рефлекс на свет, осуществляемый через II и III пару черепномозговых нервов в качестве афферентного и эфферентного путей (соответственно). Кобб утверждает, что «на сегментарном уровне в среднем, продолговатом или спинном мозге несколько нейромеров действуют .вместе как функциональная единица для того, чтобы обеспечить полезный для животного быстрый рефлекторный ответ» [145а].
Собственные волокна спинного мозга обеспечивают функциональную связь между центрами на различных уровнях и участвуют в межсегментарных рефлексах. Координация движений передних и задних конечностей, шеи и туловища, координация мышц, осуществляющих движения гв различных суставах конечности, обеспечи-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
ваются посредством таких связей. Хотя рефлексы часто рассматриваются так, как если бы их дуги были изолированы друг от друга, на самом деле между ними существует тесная связь. Облегчающие и тормозящие взаимные влияния между ними частично обусловливают описаннные выше реципрокные эффекты, а также доминирование некоторых рефлексов.
Функция внутренних органов, так же как и скелетных мышц, отчасти регулируется сегментарными рефлексами. Потоотделение, мочеиспускание, дефекация, местное сужение и расширение сосудов в ответ на охлаждение или согревание кожи — вот примеры таких рефлекторных актов. Хотя в норме эти функции зависят от импульсов, идущих от супраспинальных центров, они могут осуществляться и у спинального животного, правда, в этом случае отсутствует общая интеграция, координирующая различные виды активности, полезной для всего организма .в целом. У высших млекопитающих активность сегментарных центров в значительной степени зависит от вызывающих облегчение импульсов из других частей центральной нервной системы, особенно головного мозга. В отсутствие таких влияний рефлекторные реакции трудно, а иногда я невозможно вызвать, даже если сегментарные пути не повреждены. Аксиомой является тот факт, что специализация достигается за счет потери автономности; это отчетливо видно даже при поверхностном сравнении спинальной лягушки и человека с изолированным спинным мозгом.
В течение нескольких минут после перерезки спинного мозга лягушки в верхних сегментах ее поведение остается в основном нормальным; случайный наблюдатель не заметит ничего необычного. Напротив, человек с изолированным спинным мозгом абсолютно беспомощен, и лишь через несколько недель у него восстанавливаются рефлекторные функции, механизмы которых вовсе не были повреждены — просто их центры нуждаются в нисходящих облегчающих влияниях, от которых они зависят.
Процесс эволюции животных сопровождается выраженной эицефализацией — концентрацией нервной ткани в области головного конца. Наиболее примитивные
L
ГЛАВА II
части мозга (ствол) можно рассматривать как продолжение сегментарной системы. Но, кроме того, развивается ряд структур, которые лишены признаков сегментарное™ и которые достигают доминирующего положения чо отношению ко всем частям сегментарной системы, обеспечивая более совершенные механизмы общей интеграции в масштабе организма. Поэтому их называют надсегментарными. Наиболее примитивный из этих механизмов— ретикулярная формация, которая оказывает облегчающее и тормозящее .влияние «а различные сегментарные и межсегментарные нейроны. Уже на очень раннем этапе эволюции эта формация представляла собой сеть, объединяющую отдельные рефлекторные дуги, каждая из которых обеспечивает лишь локальную реакцию на локальное раздражение. Анатомически ретикулярная формация производит впечатление наименее специализированной нервной ткани. Это лабиринт взаимно переплетающихся волокон, между которыми довольно равномерно расположены тела клеток. Поэтому ее нельзя отнести ни к серому веществу, которое состоит главным образом из тел нейронов, ни к белому веществу, которое образовано нервными волокнами и не содержит самих клеток. Это ретикулум, или сеть, с многочисленными контактирующими между собой нейронами, имеющими короткие отростки. Деление на серое и белое вещество происходит по мере специализации функций; ретикулярную формацию можно рассматривать как основу, своего рода матрикс, для развития обоих этих веществ, с которым они сохраняют тесную связь даже у. человека. Чрезвычайно существенно, что ретикулярная формация, которая прежде всех остальных видов центральной нервной ткани получает импульсы от всех видов рецепторов на всех уровнях и образует функциональные связи со всеми эфферентными путями к эффекторам, остается хорошо развитым образованием даже у наиболее высокоспециализированных животных и сохраняет с рецепторами и эффекторами как прямые, так и непрямые связи — через скопления серого вещества и пучки волокон. В действительности ее объем, диапазон ее связей, а быть может, ,и значение возрастают, хотя до последних лет внимание исследователей было приковано
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
S?
к относительно более развитым структурам —серому веществу коры, подкорковым ядрам и проводящим путям. Еще совсем недавно на ретикулярную формацию смотрели как на остаток «сырья», из которого возникли важные части нервной системы, как на своего рода «наполнитель», не заслуживающий специального изучедия. В последнее время оценка роли ретикулярной формации в корне изменилась, и это в общем справедливо, так как на самом деле ретикулярная формация не только представляет собой материал, из которого развились более специализированные интегрирующие механизмы, но и сама является весьма важным интегратором.
Среди ядер, раньше других образовавшихся из ретикулярной формации, находятся так называемые жизненно важные центры, такие, как дыхательный я сосудо-двигательный. Они не очень отчетливо обособлены от остальной массы ретикулярной формации, но характеризуются несколько более высокой концентрацией нейронов и особым богатством связен с мускулатурой дыхательной и циркуляторной систем. Развитие многих других специализированных скоплений в ретикулярной формации у высших животных происходит параллельно с интенсивным ростом коры мозга и мозжечка.
Особенно интересно проследить развитие ладсегмеи-тарных центров, связанных с образованием специализированных органов чувств, которые сообщают чрезвычайно важную информацию организму в целом, а не отдельным его частям. Эти центры образуются на головном конце организма, т. е. в области первого контакта с меняющейся средой при движении животного. Наибольшее значение имеют дистантные рецепторы (телерецепторы), которые улавливают энергию, .выделяемую объектами, задолго до того, как животное вступает в непосредственный контакт с ними. Таковы обонятельный, зрительный и слуховой рецепторы.
Тот, кто изучает нервную систему в филогенезе, не может не поражаться огромному множеству экспериментов, которые были поставлены природой в процессе развития такого типа нервной организации, который был бы наилучшим образом приспособлен для оптимального использования информации, получаемой через специаль-
ГЛАВА If
ные рецепторы, расположенные на голове, в сочетании с информацией, получаемой от глубоких и поверхностных рецепторов тела. Каждый из специализированных органов чувств — органы равновесия, слуха, зрения и обоняния (о вкусовой чувствительности известно мало) — связан с первичной проекционной областью коры '. Эфферентные пути, идущие от этих областей к ретикулярной формации, обеспечивают возможность известной регуляции со стороны каждого из этих механизмов.
Органы равновесия в вестибулярном аппарате внутреннего уха обеспечивают рефлекторным путем ориентацию животного по отношению к горизонтальной плоскости. Кора мозжечка, получающая от них импульсы, относится к древнейшей его части, которая у человека сравнительно мала и широко заменяема по функции. В значительной мере поддержание равновесия у человека осуществляется на основе зрительной информации, и, хотя вестибулярный аппарат и древний мозжечок участвуют в регуляции позы и равновесия, это участие, как правило, не является жизненно необходимым. Следует упомянуть, что органы равновесия и древний мозжечок тесно связаны с ретикулярной формацией, а также с ядрами вестибулярного нерва, которые представляют собой скопления клеток «ретикулярного» происхождения. Хотя разрушение этого примитивного механизма равновесия у человека приводит временно к тяжелым расстройствам, постепенно происходит почти полное восстановление регуляции равновесия, осуществляемой через другие органы чувств, особенно с помощью зрения.
Примитивный слуховой мозг не достигает высокого ' уровня развития ни у одного из видов животных, хотя у птиц и рептилий, у которых слуховые области новой коры неразвиты, он является высшим слуховым ядерным образованием. У человека первичный слуховой центр представлен нижним двухолмием, размеры которого довольно малы. Через него осуществляется рефлекторное сокращение мышц среднего уха и рефлекторный поворот
* * * *
1 Подробное объяснение развития первичных коррелирующих центров читатель найдет у Эллиота [220].
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 59
глаз и головы к источнику неожиданного звука. Внезапный громкий шум вызывает старт-рефлекс, который охватывает и другие группы мышц. Однако нет основания считать, что нижнее двухолмие имеет большое значение для нервной интеграции у человека.
Примитивный зрительный мозг явно выражен уже у костистых рыб и птиц; у человека же он представлен слаборазвитым верхним двухолмием. Глаз получает огромное количество информации, и при наличии адекватного нервного механизма и эффекторов для ее использования она имеет первостепенное значение в определении поведения организма. У человека филогенетически более поздние зрительные центры, локализующиеся в коре, в значительной степени взяли на себя осуществление зрительных рефлексов и вместе с другими кортикальными областями произвольный контроль над эффекторами, участвующими сво многих из них. Например, при рефлекторной аккомодации, регулирующей толщину хрусталика, степень конвергенции глаз и диаметр зрачка при рассматривании объектов, находящихся на различном расстоянии от сетчатки, используются пути, подходящие к зрительной коре. Однако некоторые рефлексы интегрируются верхним двухолмием среднего мозга. Защитное смыкание век в ответ на внезапную вспышку света, поворот глаз и головы в сторону объектов, попадающих в периферическую часть поля зрения, зажмуривание и отдергивание головы .при внезапном приближении объекта— эти акты, очевидно, быстро осуществляются через прямые рефлекторные пути среднего мозга.
Примитивный обонятельный мозг, очевидно, имел большее значение для выживания наземных позвоночных, чем остальные рано возникшие рецепторные зоны коры. На ранних стадиях эволюции и у эмбрионов высших млекопитающих полушария образованы почти полностью обонятельным мозгом [220]. У человека и высших животных возникает массивная новая кора (неокор-текс), которая оттесняет древний обонятельный мозг. Он занимает значительно меньше места, его строение при этом изменяется, он переходит на положение второстепенной структуры соответственно той скромной роли, какую играет обонятельная информация в интеграции. Од-
ГЛАВА П,
нако обонятельный мозг составляет чрезвычайно важную часть нервного механизма, участвующего в организации эмоций, и мы тщательно рассмотрим его структуру и функции в следующей главе.
Ни одна из перечисленных нами примитивных структур не оказалась оптимальным механизмом для общей интеграции. Координация сенсорной информации в целом требовала центрального механизма. Каждый отдельный сенсорный орган обеспечивает полезную информацию, «о поведение, направляемое каким-либо одним из них, более стереотипно и в меньшей мере адаптивно, чем в том случае, если оно формируется на основе суммарной информации от всех органов чувств плюс поступающие по каналам обратной связи «текущие отчеты» от различных частей организма. Таламус — своего рода «ворота в кору» — служит центральным коллектором различных видов сенсорной информации. Расположенное рядом полосатое тело, очевидно, служило в филогенезе, до развития новой коры, основным выходом моторных сигналов из таламуса. Таким образом, таламо-стри-арную систему можно рассматривать как сенсо-мотор-нуго интегрирующую систему, хотя и примитивную, перешагнувшую в своем развитии ту стадию, на которой отдельные органы чувств функционируют как относительно независимые системы.
Наряду с участием таламо-стриарной системы в интеграции соматических функций гипоталамическая область промежуточного мозга служила (и служит) регулятором висцеральной интеграции. Ее тесную связь с лимбиче-сними структурами, в том числе обонятельными, мы рассмотрим в следующей главе, а здесь лишь укажем, что к ней идут сигналы от всех внутренних органов и что импульсы, посылаемые ею через ретикулярную формацию и связанные с нею структуры, влияют как на соматические, так и на висцеральные функции. Хотя ее основная функция состоит в регуляции гомеостаза внутренней среды, эта цель- не может быть достигнута посредством одной только вегетативной регуляции. Параллельно с основной задачей поддержания физических и химических условий, необходимых для жизни и функционирования скелетной мускулатуры, должна существовать
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 61
и способность в известной мере определять уровень этой активности. Мы увидим далее, что гипоталамус имеет важное значение для регуляции не только висцеральных, но и соматических функций.
Хотя таламус и представляет собой чрезвычайно важное образование, поведение «таламических» животных в высшей степени стереотипно и почти лишено' признаков интеллекта. Хорошим примером может служить лягушка. Большая гибкость в создании новых типов реакций «а основе обширной и самой разнообразной информации требует системы со значительно большими возможностями для избирательной активации различных комбинаций эфферентных путей. Бесчисленные преобразования и комбинации в мозгу человека возможны благодаря существованию огромного лабиринта кортикальных нейронов с их многочисленными ветвями и богатыми си-наптическими связями. Только кора, где достаточно места для широкого распространения процессов, обладает способностью к созданию многочисленных потенциальных функциональных связей, столь необходимых для обучения и проявления «разумного» поведения.
СОМАТИЧЕСКАЯ И ВЕГЕТАТИВНАЯ СИСТЕМЫ
Четкую грань между висцеральным и соматическим отделами нервной системы можно провести лишь в отношении эфферентных нервных волокон. Те из них, которые иняервируют клетки скелетных мышц, безусловно являются соматическими, а те, которые иннервируют внутренние органы, составляют вегетативную, или, по определению Лэнгли, автономную нервную систему. Термин автономный не вполне правомочен, так как хотя функции этих нервов в значительной мере независимы от волевого контроля, степень этой независимости не столь велика, чтобы можно было говорить об автономии. Так, рефлекторная реакция, возникающая при раздражении интерорецепторов, не ограничивается висцеральными сдвигами, регулируемыми вегетативными эфферентными нервами; рефлекторные реакции, вызываемые раздражением соматических рецепторов, также не ограничиваются соматическими эффекторами. Например, холод
ГЛАВА II
раздражает рецепторы кожи, возбуждение которых ведет к обширным рефлекторным висцеральным сдвигам, сопровождающимся уменьшением теплоотдачи. Один из таких сдвигов — сокращение гладких мышц в стенках кожных кровеносных сосудов, их сужение и отведение крови от кожи к глубоко расположенным областям тела, вследствие чего понижается температура кожи и уменьшается теплоотдача путем излучения. Произвольно индивидуум может перейти в более теплое место, надеть пальто или начать двигаться. Мышечная дрожь — совершенно непроизвольное повышение активности скелетных мышц — является весьма эффективным механизмом теплопродукции, который трудно отнести к чисто автономным или чисто соматическим эффектам; это результат действия импульсов, проводимых по соматическим нервам к скелетным мышцам, однако он не менее «автономен», чем любой из других, несомненно вегетативных видов активности.
Существует важное анатомическое различие между вегетативной и соматической нервной системой в способе соединения центральной нервной системы с эффекторами. Тело соматического мотонейрона расположено в пределах центральной нервной системы, а его аксон направляется к волокнам иннервируемой им скелетной мышцы. Особенность .вегетативной нервной системы состоит в том, что между центром и иннервируемым эффектором находится цепочка из двух нейронов. Тело первого лежит в головном или спинном мозге, а его аксон оканчивается в клетках вегетативных ганглиев, лежащих вне мозга; и уже аксоны этих клеток оканчиваются в области нервно-эффекторного соединения.
Хотя, как было указано вначале, вегетативная нервная система является чисто эфферентной, как правило, принято говорить о вегетативных центрах и рефлексах. При рефлекторном изменении активности висцерального эффектора эфферентным звеном служат волокна вегетативной нервной системы, и мы поэтому говорим о вегетативных рефлексах независимо от того, находятся ли соответствующие рецепторы в висцеральных или соматических структурах. Таким образом, в широком смысле слова мы можем относить к вегетативной нервной систе-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 63
ме те ее части, которые влияют на внутренние Органы. Речь идет обо всех органах, в которых эффекторы представлены гладкими мышцами или железами. Пищеварительные железы, потовые железы, слезные железы, мочевой пузырь, желудочно-кишечный тракт, сердце, кровеносные сосуды, радужная оболочка глаза иннервируются вегетативной системой. Кроме того, вегетативная система иннервирует многие железы внутренней секреции, такие, как мозговое вещество надпочечников и островки поджелудочной железы.
СИМПАТИЧЕСКАЯ И ПАРАСИМПАТИЧЕСКАЯ СИСТЕМЫ
Подразделение вегетативной системы на эти два отдела основано на анатомических и физиологических данных. Наиболее важное анатомическое различие заключается, вероятно, в локализации ганглиев. Ганглии симпатического отдела лежат, как правило, вблизи грудного и поясничного отделов спинного мозга; отсюда и название торако-люмбальная, часто прилагаемое к симпатической системе. Поскольку синапсы находятся вблизи спинного мозга, преганглионарные волокна первого эфферентного нейрона © этой двухнейронной цепи относительно коротки, тогда как постгангдионарные волокна, будучи достаточно длинными, достигают места лервно-эффекторного соединения. Парасимпатические ганглии лежат вблизи иннервируемого эффекторного органа или даже в нем самом, так что в этом случае преганглионарные волокна длинные, а постгангдионарные — короткие.
Другое анатомическое различие заключается в том, в какой области центральной нервной системы находятся клетки, дающие начало преганглионарным волокнам. Мы указали выше, что симпатическая система исходит из торако-люмбального отдела спинного мозга. Парасимпатическая складывается из двух далеко отстоящих друг от друга частей — краниальной и крестцовой, и поэтому часто говорят о кранио-сакральном отделе вегетативной системы.
Функционально симпатическая и парасимпатическая системы характеризуются антагонистическими взаимо-
ГЛАВА II
/отношениями, что также служит основой для их разгра-
|ничения.
Обозначение торако-люмбального отдела как «симпатического» происходит из отчасти воображаемой, отчасти действительно существующей тенденции симпатических разрядов к генерализации, а также из их неспецифичности; при активации одного эффектора с симпатической иннервацией .все остальные также возбуждаются как бы из «симпатии» к нему. Теперь установлено, что возможны частичные симпатические разряды [332], и все же иррадиация на несколько эффекторов более типична для симпатической системы, чем для парасимпатической. Это объясняется тем, что симпатические пре-ганглионарные волокна ветвятся и оканчиваются в ганглиях, иниервирующих различные органы, тогда как постганглионарные волокна парасимпатической системы исходят из ганглиев, лежащих непосредственно в эф-фекторном органе или на нем. Поэтому по большей части одиночное преганглионарное симпатическое волокно может действовать на ряд эффекторов, а преганглионарное парасимпатическое волокно действует лишь на один из них [296].
Двойная иннервация
Раздражение нервов, идущих к различным внутренним органам, показывает, что большинство этих органов снабжено симпатической и парасимпатической иннервацией, оказывающей противоположные влияния. Раздражение парасимпатического блуждающего нерва замедляет деятельность сердца, а симпатического — ускоряет ее. Перистальтика кишечника усиливается при раздражении парасимпатических волокон и ослабляется при раздражении симпатических. Возбуждение симпатической системы вызывает сужение сосудов; те же кровеносные сосуды, которые иннервируются парасимпатическими нервами (в немногих ограниченных областях), расширяются при их активации.
Из приведенных выше примеров ясно, что ни симпатическую, ни парасимпатическую систему в целом нельзя классифицировать либо как тормозную, либо как воз-
;ПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
буждающую, но любой орган, иннервируемый обеими системами, возбуждается одной из них и тормозится другой.
Механизм антагонистических взаимоотношений между симпатической и парасимпатической системами
Противоположное действие двух отделов вегетативной нервной системы на эффекторы объясняется тем,- что химические медиаторы, выделяющиеся на месте нервпо-эффекторного контакта, различны, .Парасимпатический медиатор — ацетилхолин, а симпатический — норадрена-лин (с некоторыми исключениями). Соответственно передачу возбуждения называют холинэргической и ад-ренэргической. По большей части висцеральные эффекторы, возбуждаемые ацетилхолином, тормозятся норад-реналином, и наоборот.
Объяснение периферических влияний симпатической и парасимпатической систем не раскрывает механизма центрального распределения импульсов на эфферентные нейроны двух противоположных систем '. Это .имеет важнейшее значение при анализе причин и лечении эмоциональных и психических расстройств, в которых вегетативная система играет важную роль. Психотерапевт должен уметь воздействовать на центральную нервную систему путем .внушения и беседы. Афферентный приток частично изменяется под влиянием того, что он говорит, и тех условий, в которые он помещает больного. Это приводит к изменению характера центральной активности, к изменению поведения больного и, таким образом, к изменению информации, идущей по обратным связям от его эффекторов, и к преобразованию его взаимодействия со средой. Если, кроме того, используются химио-терапевтические средства, то возможности регуляции центральных процессов значительно расширяются, так как область химических влияний и их диапазон практи-* * * *
1 Существуют некоторые исключения из общего правила об их антагонистическом действии, но мы не будем их здесь рассматривать.
3-517
ГЛАВА II
чески неограниченны. Ниже мы уделим особое внимание анализу влияния различных веществ и других терапевтических воздействий на вегетативные функции и поведение, а также рассмотрению возможных механизмов этих влияний.
В норме изменения вегетативной активности могут быть вызваны различными способами. Повышая и понижая активность некоторых афферентных нервов, можно изменять вегетативные рефлексы, влияющие на сердце, кровеносные сосуды, кишечник и другие внутренние органы. Эти рефлексы сохраняются при глубоком наркозе и после удаления части мозга или всего мозга в целом. Они могут исходить от любых рецепторов, в том числе от расположенных во внутренних органах. Например, болевые раздражения, возникающие при растяжении кишечника или действии тепла на кожу, вызывают изменение частоты сердечных сокращений и сопровождаются рефлекторным изменением просвета кровеносных сосудов. Это в свою очередь приводит к изменению уровня артериального давления. Подобные изменения можно вызвать в эксперименте раздражением некоторых областей центральной нервной системы.
Центральные и периферические вегетативные сдвиги— неизбежные сьушики эмоциональных расстройств. Наша ближайшая задача состоит в' описании функций центров, которые регулируют поток вегетативных импульсов к внутренним органам; это позволит нам подготовить читателя к обсуждению связи между эмоциями и активностью вегетативной нервной системы.
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ СПИННОГО МОЗГА
Симпатические и парасимпатические нервы находятся в состоянии постоянной активности, или тонуса, даже в отсутствие внешних раздражений. Этот тонус поддерживается активностью вегетативных центров, локализующихся главным образом в продолговатом мозге и гипоталамусе, но имеющихся и з спинном мозге. Последние активируются афферентными импульсами, идущими к ним через афферентные нейроны от рецепторов, а также от продолговатого мозга и гипоталамуса. В отсутствие
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 67
супраспинальных влияний (например, после пересечения нисходящих путей) спинномозговые вегетативные рефлексы осуществляются автономно. Так, раздражение седалищного нерва вызывает повышение частоты сердечных сокращений и сужение сосудов, особенно в области живота. Такие рефлексы осуществимы даже у спиналь-ного животного, у которого путем поперечной перерезки спинной мозг отъединен от головного. У интактного животного наблюдаются сходные, но более генерализованные и более эффективные сдвиги.
Изменение внутренней среды влияет «а вегетативную активность сшшального животного. Гипогликемия вызывает симпато-адреналовый разряд, а гиперкапния и асфиксия приводят к повышению кровяного давления [16], но чувствительность к этим раздражителям у такого животного значительно ниже, че.м в норме.
Прямое раздражение спинальных центров приводит к частичной активации симпатической системы, причем характер реакции зависит от места раздражения [332]. Например, если электроды находятся в торако-люмбаль-ной части спинного мозга, даже самое незначительное по силе раздражение вызывает резкое повышение кровяного давления. Аналогичное раздражение в области нижнего шейного отдела спинного мозга вызывает симпатическую реакцию со стороны глаза (расширение зрачка, сокращение третьего века), но не приводит к повышению кровяного давления. Это лишь один пример того, что симпатическая система не всегда действует как единое целое, так что истинное значение этой системы нельзя оценить без учета ее способности к локальному, или частичному, возбуждению. Частичные симпатические разряды также возможны при раздражении продолговатого мозга и гипоталамуса.
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ ПРОДОЛГОВАТОГО МОЗГА
Жизненно важные симпатические и парасимпатические центры находятся в продолговатом мозге, где они четко локализованы. Электрическое раздражение симпатических центров продолговатого мозга у экспериментальных животных вызывает отчетливые реакции во
3*
68 ГЛАВА II
всех областях тела. Они выражаются в повышении частоты сердечных сокращений, повышении кровяного давления, максимальном расширении зрачка, сокращении третьего века, пилоэрекции, потоотделении, ослаблении перистальтики кишечника и повышении содержания сахара в крови. В основе повышения содержания сахара в крови лежит следующий механизм: симпатическая активация мозгового вещества надпочечников приводит к выделению активных гормонов, которые стимулируют процесс распада гликогена в печени. Мозговое вещество выделяет адреналин и норадреналин, о которых мы подробно скажем ниже при обсуждении физиологии эмоций.
Активность симпатических центров продолговатого мозга в равной степени повышается в ответ на рефлекторное или химическое раздражение. При вдыхании воздуха с повышенным содержанием СОг или пониженным содержанием Ог у интактного животного возникают характерные симптомы возбуждения симпатической системы. Асфиксия при пережатии трахеи вызывает мощный разряд в симпатических центрах в результате комбинированного влияния гиперкапнии и гипоксии. После высокой перерезки спинного мозга та же степень асфиксии весьма незначительно сказывается на функции органов, имеющих симпатическую иннервацию; отсюда ясно, что указанные реакции почти целиком опосредуются центрами, лежащими выше спинного мозга. Установлено, что С02 оказывает прямое раздражающее действие на симпатические центры продолговатого мозга [351]; понижение содержания 02 выражается в прямом подавлении их возбудимости. Однако, если напряжение кислорода в жидкостях организма падает очень низко, возбуждаются хеморецепторы каротидного синуса, что приводит к рефлекторной активации симпатических центров продолговатого мозга, несмотря на то, что их возбудимость в условиях гипоксии понижена.
Тщательными исследованиями установлено, что симпатический центр (пресс'орная зона) расположен в основном в ростральной части продолговатого мозга [16]. Перерезка в этой области приводит к резкому падению кровяного давления и исчезновению тонических импульсов, регистрируемых в ускоряющем нерве сердца. Де-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 69
прессорная зона расположена в каудальной части продолговатого мозга и также характеризуется тонической активностью. Расширение сосудов, вызываемое раздражением депрессорной зоны, а также усилением разрядов барорецепторов, связано не с активацией парасимпатических сосудорасширяющих нервов, а с торможением симпатического центра. Вот почему у симпатэктомиро-ванного животного сосудорасширяющие эффекты отсутствуют '.
Прямое раздражение центра замедления сердечной деятельности (тесно связанного с ядром блуждающего нерва) или его рефлекторное возбуждение приводит к падению кровяного давления, зависящему от замедления сердечного ритма. При этом активность желудочно-кишечного тракта повышается2.
Следует подчеркнуть, что центр блуждающего нерва и депрессорная зона, подобно симпатическому центру продолговатого мозга, постоянно активны, а поскольку их действия противоположны, это обеспечивает контроль и регуляцию кровяного давления. Сосудосуживающий центр способствует поддержанию нормального тонуса кровеносных сосудов посредством импульсов, идущих по симпатическим волокнам к гладкой мускулатуре их стенок. Депрессорная зона, вероятно, понижает активность сосудосуживающего центра (прессорная зона) посредством реципрокного торможения. Центр ускорения сердечной деятельности (симпатический) повышает возбудимость водителя ритма сердца, а центр, тормозящий сердце (парасимпатический), противодействует этому влиянию. Таким образом, устранение любого из этих центров приводит к снятию его тонического влияния и доминированию антагонистической системы. * * * *
1 Расширение сосудов скелетных мышц, вызываемое раздражением гипоталамуса, не зависит от целостности депрессорной зоны продолговатого мозга, а связано с активацией холинэргических симпатических нервов [900а].
2 Среди других парасимпатических реакций, вызываемых раздражением продолговатого мозга, следует упомянуть рефлекторную рвоту. Она возникает при раздражении различных афферентных нервов, а также при изменениях внутренней среды. В этой реакции участвует продолговатый мозг [80],
\
70 ГЛАВА II
ВЕГЕТАТИВНЫЕ ЦЕНТРЫ ГИПОТАЛАМУСА И ИХ ФУНКЦИИ
Раздражение гипоталамуса
Гипоталамус — это совсем маленький участок мозга, но вряд ли существует какой-либо другой орган, Который имел бы столь важное значение для благополучия всего организма в целом. Он лежит в основании мозга, за перекрестом зрительных нервов, над «главной железой» организма — гипофизом, с которым ои тесно связан анатомически и функционально (см. [692, 736]).
В гипоталамусе, как и в продолговатом мозге, симпатические и парасимпатические эффекты вызываются из различных участков, хотя здесь это разделение не столь четко выражено. Парасимпатические реакции возникают при раздражении передней области (непосредственно за хиазмой), а симпатические — при раздражении задней и латеральной областей. Так, электрическое раздражение заднего гипоталамуса приводит к сужению сосудов брюшной полости и повышению кровяного давления. Кроме того, оно вызывает расширение зрачка, •сокращение третьего века и пилоэрекцию, но подавляет перистальтику кишечника.
Весьма интересно, что достаточно интенсивное раздражение симпатических центров гипоталамуса может привести к повышению секреции гормонов мозгового вещества надпочечников, влияние которых складывается с действием общего разряда симпатической системы. Это отчетливо видно по реакции органа, лишенного симпатической иннервации. Из ф,нг. 2, Б видно, что слабое раздражение гипоталамуса вызывает сокращение интактио-го (/), но не денервированного (//) третьего века, незначительное повышение кровяного давления и умеренное учащение сердечного ритма. При более интенсивном раздражении (ф.иг. 2, А) не только усиливаются эти симпатические реакции, но и происходит сокращение денервированного третьего века. Поскольку возможность его возбуждения нервным путем исключена (так как оно денсрвировано), можно предположить, что возбуждающим агентом служит химическое вещество, приносимое кровью. Соответствующие эксперименты показали, что
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ 71
речь идет о гормоне мозгового вещества надпочечников. Этот гормональный эффект проявляется значительно позже, чем прямой эффект возбуждения симпатического нерва. Такая задержка характерна для гормональных влияний и связана с тем, что для переноса гормонов от соответствующих желез до органов-мишеней необходимо некоторое время [316].
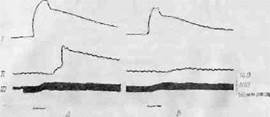
Фиг. 2, Нервные и гуморальные симпатич?ские влияния при раздражении заднего отдела гипоталамуса [316].
Опыты па кошке под пентоталовым наркозом н флакееднлом. Характеристика раздражений: А. 1,5 в, 71 имп/сек. продолжительность импульса 1,6 мсек, длительность раздражения 7 сек. 15. Те же условий, но продолжительность импульса 0,8 мсек. Видно, что более длительные импульсы вызывают секреторную активность мозгового вещества надпочечников, что ведет к сокращению денервированного третьего века. / — нормальное третье веко; // — де-исринрованнос третье веко; /// -— кровяное давление.
Генерализованный характер реакции, вызываемой раздражением гипоталамуса, иллюстрирует фиг. 3, на которой приведена запись активности мочевого пузыря, кишечника, денервированного третьего века и кровяного давления. Раздражение заднего гипоталамуса вызывает ослабление перистальтики кишечника и сокращение мочевого пузыря, повышение кровяного давления и секреторную активность мозгового вещества надпочечников (о чем свидетельствует сокращение денервированного третьего века). Все эти реакции совпадают с результатами раздражения эфферентных симпатических нервов, идущих к соответствующим органам. Следует отметить, что на кишечник и мочевой пузырь симпатическая
η
ГЛАВА li
система оказывает тормозящее' влияние, а на гладкие мышцы кровеносных сосудов и миокард (не регистрировалось) — возбуждающее. Данный эксперимент показывает, что гипоталамус может быть источником генерализованного симпатического разряда, подавляющего одни вегетативные эффекторы и возбуждающего другие [318].

Фиг. 'J. Широко иррадирующая симпатическая и симпато-адренало-
вая реакция на слабое раздражение заднего отдела гипоталамуса
[318].
Характеристика раздражения: 1 в,'99 имп/сек, 0,8 мсек, 15 сек. / — мочевой
пузырь; // — двенадцатиперстная кишка; /// — кровяное давление; IV— де-
нервировапное третье веко.
Как упоминалось выше, парасимпатические реакции легче вызвать путем раздражения переднего гипоталамуса. Такое раздражение повышает тонус и усиливает перистальтику кишечника, снижает кровяное давление и частоту сердечных сокращений, т. е. производит действие, прямо противоположное только что описанному 1- * * * -,
1 Тормозящее влияние на мочевой пузырь зависит от секреции мозгового вещества надпочечников.
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
(при раздражении симпатического центра заднего гипоталамуса повышается кровяное давление и снижается гонус кишечника). При раздражении переднего (парасимпатического) отдела гипоталамуса происходит сокращение мочевого пузыря (органа, имеющего парасимпатическую иннервацию) и сужение зрачка.
Хотя, раздражая различные отделы гипоталамуса, можно разделить симпатические и парасимпатические эффекты, не исключена возможность смешанных реакций. Указанные зоны гипоталамуса в какой-то степени перекрываются, что особенно убедительно показал в своих экспериментах Гесс [454]. Кроме того, при раздражении заднего гипоталамуса наряду с симпатическими центрами возбуждаются волокна, идущие от расположенных рострально парасимпатических центров. В результате этого возникают комбинированные симпатические и парасимпатические реакции, а поскольку обычно они являются антагонистическими, конечный эффект для определенного органа может быть незначительным или нулевым. Существенное значение имеет не только локализация, но и характер раздражения. Например, раздражение с низкой частотой обычно вызывает парасимпатические реакции, а раздражение той же точки с высокой частотой даст симпатический эффект. Наконец, следует иметь .в виду, что периферические реакции, вызываемые раздражением гипоталамуса (например, повышение кровяного давления), нередко влекут за собой вторичные вегетативные рефлексы, которые видоизменяют первичный результат раздражения гипоталамуса.
Местная анестезия и разрушение областей гипоталамуса
Обсуждавшиеся до сих пор факты касаются фазиче-ских, или кратковременных, реакций на раздражение. А между тем гипоталамус, подобно продолговатому мозгу, оказывает тонические, или постоянные, влияния на вегетативную систему. Их легко продемонстрировать, исключив иа время или навсегда влияние части переднего или заднего гипоталамуса. Сделать это можно либо путем введения небольших количеств барбитуратов
74 ГЛАВА II
в специфические области гипоталамуса (временная блокада), либо путем очагового разрушения токами высокой частоты (стойкое разрушение).
Если задний гипоталамус действительно оказывает тонические симпатические влияния, то его разрушение или снижение его активности местным введением анестезирующего вещества должно привести к падению кровяного давления и снижению частоты сердечных сокращений. Эксперименты показали, что это действительно так. Аналогичные воздействия на область переднего гипоталамуса, исключающие или снижающие парасимпатические разряды, должны были привести к повышению кровяного давления и частоты сердечных сокращений. Это также удалось получить в эксперименте, хотя эффект был не столь демонстративен, как в опытах по исключению симпатической активности заднего гипоталамуса. Можно сделать вывод, что гипоталамус обладает тонической активностью как в отношении симпатической, так и .в отношении парасимпатической системы, но что $ норме симпатический тонус на этом уровне значительно выше парасимпатического [356, 741].
Соматические эффекты при раздражении гипоталамуса
При изучении поведения экспериментальных животных в состоянии бодрствования или поверхностного наркоза было отмечено, что раздражение гипоталамуса вызывает не только вегетативные, по и соматические реакции. Симпатические разряды, вызываемые раздражением заднего гипоталамуса, сопровождались изменениями полы - кошка выгибала спину, выпускала когти, дыхание ее заметно учащалось; она пыталась укусить экспериментатора. Весь вид кошки с расширенными зрачками, взъерошенной шерстью на спине и хвосте олицетворял ярость. Возбудимость всей соматической системы в целом была повышена.
Эта картина прямо противоположна той, которая возникает при раздражении переднего гипоталамуса. Парасимпатические реакции, такие, как сужение зрачков, падение кровяного давления и уменьшение частоты сер-
ПРИНЦИПЫ ОРГАНИЗАЦИИ НЕРВНОЙ СИСТЕМЫ
дечных сокращений, сопровождаю юн расслаблением скелетных мышц, и животное засыпает. В других экспериментах раздражение переднего гипоталамуса вызывало мочеиспускание и дефекацию с характерными позами. Приведенные опыты показывают, что вегетативные реакции, возникающие при раздражении гипоталамуса, связаны с целым комплексом соматических — симпатические реакции сопровождаются повышением возбудимости соматической нервной системы, парасимпатические характеризуются параллельным снижением возбудимости соматической нервной системы [453].
Наконец, следует упомянуть, что изменения ритма и глубины дыхания также могут быть .вызваны раздражением гипоталамуса. Обычно повышенная распиратор-ная активность связана с симпатическими влияниями, а пониженная— с парасимпатическими [454]. Повреждение заднего гипоталамуса приводит к снижению легочной вентиляции [743].
Вегетативная функция коры
Симпатические и парасимпатические реакции могут быть вызваны раздражением коры или различных частей лимбической системы. Раздражение коры вызывает в целом более слабый и не столь генерализованный эффект, какой возникает при раздражении гипоталамуса или продолговатого мозга. Некоторые вегетативные реакции устраняются после перерезки пирамидного тракта, другие передаются через гипоталамус к низшим вегетативным центрам. Вызванные раздражением коры прессор-ные реакции связаны с сужением сосудов почек; если почка денервирована, эти реакции не возникают. Удаление моторной коры сопровождается сужением кожных сосудов противоположной стороны тела. Это показывает, что тоническая иннервация в физиологических условиях поддерживается корой. Недавно было показано, что при раздражении моторной коры (а также гипоталамуса) повышается кровоснабжение задних конечностей. Дальнейшими исследованиями установлено, что расширение наблюдается только в сосудах мышц и зависит от хо-линэргических вегетативных нервов, так как оно спи-
ГЛАВА И
мается атропином и сохраняется при перевязке спинного мозга в поясничном отделе, когда импульсы от мозга могут достигнуть этих мышц только через симпатическую цепочку [217]. Этим расширением сосудов объясняется повышение кровотока, связанное с началом произвольного сокращения мышц (а также, возможно, с тем, которое возникает, когда мы думаем о движении, не производя его в действительности).
Это явление служит примером того, что вызванные раздражением коры вегетативные разряды связаны с соматическими функциями коры. Двигательная активность коры сопровождается расширением сосудов в мышцах и повышением секреторной активности потовых желез в области ладоней (считается, что пот способствует удержанию предметов в руках) [164]. Раздражение зрительной коры сопровождается изменением диаметра зрачка, а акт жевания, связанный с раздражением коры, сопровождается слюноотделением. Напротив, состояния, сопровождающиеся возбуждением гипоталамуса или продолговатого мозга, приводят к генерализованной симпатической реакции.
ГЛАВА
НЕРВНЫЙ СУБСТРАТ ЭМОЦИЙ
Название этой главы отражает наше глубокое убеждение в том, что в основе эмоций, как ощущаемых, так и выражаемых, лежит активность нервной системы. Путем изменения активности желез и мышц в эмоциональную деятельность вовлекается весь организм, но основой эмоций все же служит центральная нервная система.
Возникает вопрос, не принимают ли какие-то определенные части центральной нервной системы более прямого участия в эмоциональных процессах по сравнению с другими, и если да, то каким образом. На протяжении всего изложения перед нами отчетливо выступает первостепенная роль гипоталамуса, это убеждает нас в том, что ему принадлежит такое же место и в механизме интеграции эмоций. Его значение объясняется не тем, что он является «седалищем» эмоций, или каким-то «эмоциональным центром», или группой таких центров. Скорее оно связано с его исключительными взаимоотношениями с остальными отделами нервной системы. Это небольшая, но сложная структура, связи и взаимодействия которой с другими отделами нервной (и эндокринной) системы имеют особое значение для состояний организма, лежащих в основе эмоций.
КОРТИКО-ГИПОТАЛАМО-ЛИМБИЧЕСКИЕ ВЗАИМОДЕЙСТВИЯ
Давно известно, что выражение эмоций может осуществляться в отсутствие новой коры. Однако активность новой коры, несомненно, имеет важное значение для интеграции эмоциональных ощущений. Брэйди [90]
глАвА Ш
дал краткий обзор первых клинических и экспериментальных данных, касающихся участия коры в эмоциональных процессах. Эти данные указывают на особую роль лобных долей. Точно установлено, что повреждения лобной и префронтальной областей или их удаление снижают эмоциональную реактивность, однако довольно часто такие разрушения приводят к повышенной эмоциональной лабильности, которая характеризуется резкими колебаниями эмоционального состояния. Сдвиги, возникающие в результате участия коры, должны рассматриваться в плане интегративных отношений между лим-бической системой и диэнцефало-кортикальными взаимодействиями.
Следует подчеркнуть, что, как показали Мерфи и Гельгорн [671], раздражение гипоталамуса оказывает сильное генерализованное облегчающее влияние на активность новой коры. Такое облегчение наблюдается при разных условиях. Вызванные раздражением коры сокращения мышц и разряды в пирамидных путях усиливаются при одновременном раздражении гипоталамуса [310]. Раздражение гипоталамуса подавляет «диа-ловые» волны в электрокортикограмме, приводит к повышению частоты и амплитуды кортикальных воли, а также частоты стрихнинных пиков, вызываемых местным нанесением стрихнина на кору. Таким образом, раздражение гипоталамуса при самых разнообразных условиях приводит к повышению активности новой коры. Это возбуждение коры бывает двухсторонним, хотя оно сильнее выражено на ипсилатаральной стороне. Оно было обнаружено в моторной, сенсорных и ассоциативных зонах, а также в базальных ядрах мозга. Обширность кортикальных зон, доступных облегчающим влияниям со стороны гипоталамуса, указывает на необходимость их учета при анализе интегративного действия нервной системы, в том числе функций коры. Без всестороннего учета влияний гипоталамуса нельзя изучать механизмы, лежащие в основе деятельности и эмоций как в норме, так и в патологии.
В другой статье Мерфи и Гельгорна [672] показано, что, поскольку гипоталамус способен активировать ассоциативные области коры, он, по-видимому, им
Дата добавления: 2014-12-10; просмотров: 1408;