Установление строения и структуры молекул ДНК и РНК
Важнейшие полимеры — молекулы ДНК и РНК — состоят из мономеров, называемых нуклеотидами. Как белки состоят из последовательно соединенных аминокислот, так и нуклеиновые кислоты — из последовательно связанных между собой нуклеотидов.
Эти полинуклеотиды впервые выделил швейцарский врач Ф.Мишер (1868); он назвал это вещество нуклеином, затем немецкий химик Р. Альтман предложил название — нуклеиновая кислота, так как это вещество проявляет кислотные свойства, обнаруживаемые преимущественно в клеточном ядре. Мишер не знал, какое важное открытие он сделал, но писал об этом так: «Обрабатывая клетки гноя слабыми щелочными растворами, я получил в результате нейтрализации раствора осадок, который не растворялся ни в воде, ни в уксусной кислоте, ни в разведенной соляной кислоте, ни в обычном солевом растворе и который не мог принадлежать ни к одному из белков, известных в настоящее время».
Сотрудник Мишера А. Коссель обнаружил, что в состав нуклеиновых кислот входят пуриновые (А, Г) и пиримидиновые основания (Ц, Т) и простейшие углеводы; он выделил аденин (А) и гуанин (Г), фосфорную кислоту и углеводы. Если в построении белка участвует 20 аминокислот, то нуклеотидов — всего 4 (хотя сами они — достаточно сложные образования). К началу 1900 г. в лаборатории П.Левина (США) расшифровали углеводную часть кислот. У всех живых существ молекулы ДНК и РНК построены по одному плану, каждый нуклеотид состоит их трех компонентов, соединенных химическими связями: из одной молекулы фосфорной кислоты, одной молекулы сахара и одной молекулы органического основания. Их фосфатные группы освобождают в растворах ионы водорода. Сахар может быть в двух вариантах: рибоза (Р), представляющая сахар с пятью атомами углерода, к одному




из которых присоединена гидроксильная группа (—ОН), и дезок-сирибоза (Д), в молекуле которой в отличие от глюкозы не 6, а 5 атомов углерода (пентоза) и к одному из атомов углерода присоединен атом водорода. При этом они никогда не встречаются одновременно, поэтому этим сахарам соответствуют два типа нуклеиновых кислот — ДНК и РНК. Сначала думали, что они тоже разобщены в клетках: ДНК — в ядре, а РНК — вне его. Теперь ясно, что ДНК находится в основном в ядре (хромосомах), а частично — в других клеточных компонентах (например, хлоропла-стах зеленых растений). РНК содержится как в цитоплазме, так и в ядрышке. Кроме того, иногда временно цепь ДНК соединяется с цепью РНК.
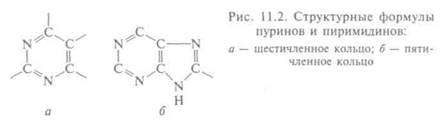
Основания — другой компонент нуклеотида — названы так, потому что реагируют как основания: в кислой среде способны присоединять ион водорода. Они тоже могут относиться к двум группам: пиримидинов, в основе строения которых — шестичлен-ное кольцо (рис. 11.2, а), и пиринов, у которых к пиримидиново-му присоединено пятичленное кольцо (рис. 11.2, б). В ДНК входят два пурина (А, Г) и два пиримидина (Ц, Т), а в РНК — только три: А, Г и Ц, а вместо тимина — другой пиримидин — урацил (У), т.е. в составе каждой из нуклеиновых кислот находится по 4 основания. В ДНК последовательно соединены дезоксирибонуклеотиды, каждый из которых содержит какое-то из четырех оснований (А, Ц, Г, Т), а РНК — рибонуклеотиды, содержащие тоже по одному основанию (А, Ц, Г, У). Все молекулы имеют форму цепи (от 77 до нескольких миллионов нуклеотидов).
У.Астбери, автор термина «молекулярная биология», вместе с Ф. Беллом получил рентгенограммы ДНК (1938). Они показали, что азотистые основания располагаются одно за другим, построенные как пластинки. В 1948 г. английский химик-органик А.Тодд, подробно изучая структуру ДНК, выяснил, как связываются между собой четыре азотистых основания с пятиатомным кольцом сахара рибозы или дезоксирибозы и молекулой фосфорной кислоты. Нуклеотиды — не только составная часть нуклеиновых кислот, они входят в состав ферментов в качестве активных групп — коферментов (так Тодд назвал комплекс азотного основания, углевода и остатка фосфорной кислоты). Блоки А, Г, Т, Ц образуют длинную полимерную цепь, соединяясь друг с другом в разных комбинациях.
Американский биохимик Э.Чаргафф сформулировал (1948) правила регулярности в парных отношениях пуриновых и пиримидиновых оснований в молекулах нуклеиновых кислот: 1 — общее количество гуанина и аденина (из группы пуринов Г и А) равно количеству цитизина и тимина (из группы пиримидинов Ц и Т), т.е. А + Г = Т + Ц; 2 — отношения А/Т и Г/Ц примерно равны единице, т.е. А = Т и Ц = Г; 3) — при этом Г + Т = А + Ц; 4 — ДНК из разных источников может иметь отличия — в одних случаях А + Т>Г + Ц, ав других — Г + Ц>А + Т. Эти правила явились предтечей открытия двойной спирали ДНК.
Для молекулы ДНК тоже характерна структура трех видов — первичная, вторичная и третичная. Первичная структура ДНК состоит из нуклеотидных цепей, у которых скелетную основу составляют чередующиеся сахарные и фосфатные группы, соединенные ковалентными связями, а боковые части представлены одним из четырех оснований и присоединяются одна к другой молекулой сахара. Нуклеотиды расположены друг за другом и связаны ковалентно с фосфатом и сахарным остатком, образуя по-линуклеотидную цепь.
Вторичная структура была сформулирована Д.Уотсоном и Ф. Криком. Две идущие рядом нити, скрепленные одна с другой перемычками и свившиеся в двойную спираль, и есть молекула ДНК. Обе нити одинаковы по длине, остатки пар А—Т и Г—Ц разделены одинаковыми расстояниями. Двойная спираль имеет упорядоченный характер, так как каждая связь основание — сахар находится на одинаковом расстоянии от оси спирали и повернута на 36°, причем в каждой из них в зависимости от вида ДНК могут быть до миллионов блоков — нуклеотидов. Порядок их чередования определяет наследственную информацию, записанную в ДНК и передаваемую следующим поколениям. Первое предположение о роли нуклеиновых кислот в качестве генетического материала сформулировал доцент Петербургского университета А. Щепотьев (1914). Химики понимали, что ДНК собрана из нуклеотидов, имеющих фосфатную группу, связанную ковалентно с пятиуглеродным сахаром, который связан с одним из четырех азотистых оснований. Нуклеотиды соединены друг с другом так, чтобы фосфатная группа одного была связана с сахаром предыдущего, и из их чередующихся комбинаций образуется длинная цепочка — сахарофосфатный остов молекулы. По одну сторону под прямым углом к остову располагаются основания.
Молекула ДНК оказалась закручена в спираль: снаружи спирали — остов, а внутри — перпендикулярные ему основания. На один виток спирали приходилось примерно по десять нуклеотидов, а ее толщина указывала, что скручено более одной нити. Итак, вторичная структура отражает форму нуклеиновой кислоты. Степень скручивания ДНК зависит от ферментов.
Л
Р.Франклин исследовала на фотопленке пятна от рентгеновского излучения, рассеянного кристаллами очищенной ДНК (1952). В обсуждении результатов принимал активное участие и физик М.Уилкинс, работавший в той же лаборатории. Полученные рентгенограммы стимулировали многих ученых к поиску модели структуры ДНК. История открытия структуры ДНК описана американским биохимиком Дж. Уотсоном в его книге «Двойная спираль» (1968). В 1951 г. Уотсон встретился в Копенгагене с Уилкинсом и ознакомился с рентгенограммами ДНК. Руководитель Уилкинса Сальвадор-Лурия договорился о стажировке Уотсона в Кавендишской лаборатории, где работала группа ученых, занимавшихся рен-тгеноструктурным анализом сложных биомолекул и сравнивавших свои модели с опытными данными, полученными на первых, весьма несовершенных ЭВМ. В Кембридже Уотсон познакомился с Криком, физиком, переквалифицировавшимся в биохимика, и узнал, что структурные формулы химиков были далеки от совершенства.
Уотсон и Крик, разобравшись в структуре пуринов (А, Г) и пиримидинов (Т, Ц), решили, что они должны быть связаны между собой. Для объяснения правила Чаргаффа ДНК должна состоять из двух цепей, которые должны закручиваться так, чтобы сохранялись определенные углы между разными группами атомов. И возникла двойная спираль, в которой пурины и пиримидины выстроены по типу ступенек лестницы: «перекладины» — основания, «веревки» — сахарофосфатные остовы.
Каждая перекладина образована из двух оснований: А и Т или Г и Ц, что и объясняет правило Чаргаффа. Так как в каждой паре есть одно основание с одним кольцом и одно — с двумя, величина перекладин одинаковая, и остовы цепей находятся на одном расстоянии. Основания присоединены к двум противоположным цепям, удерживаемым водородными связями между основаниями. Поскольку звеньями цепи являются пары Ц с Г и А с Т, удобнее использовать образ лестницы, составленной из ступенек-пар ЦГ, ГЦ, ТА и AT, следующих в определенном порядке. Из-за закрученности в спираль молекула похожа на винтовую лестницу со ступеньками из пар нуклеотидов.
В живых клетках цепи очень длинные, содержат до 108 пар в ряд и свиты в плотный клубок. У человека длина такой винтовой лестницы в размотанном состоянии достигает нескольких метров, и это одна молекула! Отсюда — огромность числа возможных вариантов расположения молекул в ДНК. И это разнообразие связано с разнообразием жизни, а расположение четырех типов пар в молекуле ДНК задает всю программу, говорит клетке, как ей развиваться и что делать.
Диаметр двойной спирали 2 • 10-9 м (2 нм), расстояние между соседними парами оснований спирали 0,34 • 10-9 м (0,34 нм), полный оборот спирали завершается через 10 пар, а длина зависит от организма, которому принадлежит эта молекула ДНК. Длина плодовой мушки (дрозофилы) 4 • 10-3 м, а самой длинной ее хромосомы — в 10 раз больше. У простейших вирусов ДНК содержит несколько тысяч звеньев, у бактерий — несколько миллионов, а у высших — миллиарды. Если выстроить в одну линию молекулы
ДНК, заключенные в одной клетке человека, то получится длина в 2 м, т. е. длина в миллиард раз больше толщины. Но она умещается в клеточном ядре, значит, ее «укладка» такова, чтобы по всей длине она была доступна для белков, которым нужно «читать» гены. Основания, соединенные слабой водородной связью, взаимно дополняют друг друга, и каждая цепь автоматически поставляет информацию для нахождения партнера. В эукариотичес-ких клетках основные части ДНК и белков сплетены так, что напоминают нить бус. Каждая такая «бусинка» окружена четырьмя ядерными блоками и содержит около 200 сдвоенных оснований, а «нить» состоит из ДНК и ядерного белка (гистона), отличного от того, что входил в состав «бусинок».
О расшифровке структуры ДНК сообщалось в статье Уотсона и Крика, занявшей всего две странички в журнале, но открывшей новую эпоху в раскрытии тайны жизни. В публикации (1953) Крик и Уотсон отметили, что такая структура хорошо объясняет и процесс «воспроизводства» этой молекулы. При рассоединении цепей возможно присоединение новых нуклеотидов к каждой из них, тогда около каждой старой возникнет новая цепь, точно ей соответствующая. Так впервые пришли к структуре, способной к самовоспроизведению. Число два удовлетворило биологов, поскольку и клетки, и хромосомы воспроизводятся путем деления исходной на две.
Третичная структура ДНК, определяемая трехмерной пространственной конфигурацией молекул, пока изучена недостаточно.
Исследования показали, что ДНК может существовать в двух формах: А (при низкой влажности) и В (при высокой). Для обеих форм построили молекулярные модели. Из дифракционных картин волокон ДНК информацию получить было достаточно трудно, так как у цепи ДНК вдоль оси расположены волокна беспорядочно, но была подтверждена ее спиральная структура. К настоящему времени исследователи научились синтезировать в необходимом количестве и получать в достаточно чистом виде короткие участки ДНК заданной последовательности, что позволяет закристаллизовать фрагменты молекулы длиной от 4 до 24 пар оснований и исследовать эти кристаллы с помощью рентгеноструктурного анализа. Исследования дали действительную похожесть обеих форм на гибкую лестницу, закрученную спирально вокруг центральной оси.
Дата добавления: 2014-12-27; просмотров: 1743;