Различают следующие уровни организации биологических систем.
| Стр. | |
| Пояснительная записка……………………………………………... | |
| ПРИМЕРНЫЙ ТЕМАТИЧЕСКИЙ ПЛАН………………………………….. | |
| Содержание учебноГО МАТЕРИАЛА …...………………………….. | |
| Общая эпидемиология……………………………………………………… | |
| Иммунопрофилактика инфекционных болезней……………………………. | |
| Частная эпидемиология…………………………………………………….. | |
| Внутрибольничные инфекции………………………………………………... | |
| ИНФОРМАЦИОННО-МЕТОДИЧЕСКАЯ ЧАСТЬ………………………… | |
| Литература………………………………………………………………..…… | |
| Рекомендуемые средства диагностики……………………………………… | |
| Перечень рекомендуемых наглядных пособий………………………...…… | |
| Краткие методические указания по использованию персонального компьютера, технических средств обучения………………………………... |

[1] Иллюстрации по каждой нозологической форме, отражающие интенсивность, динамику, структуру и территориальное распространение заболеваемости.
Различают следующие уровни организации биологических систем.
1. Молекулярно-генетический (элементарными структурами служат коды наследственной информации, которые передаются из поколения в поколение, элементарными явлениями — воспроизведение этих кодов).
2. Онтогенетический (элементарными структурами служат клетки, элементарными явлениями — их деление).
3. Популяционно-эволюционный (элементарными структурами являются популяции, элементарными явлениями — изменение генофонда популяций, которые приводят к возникновению приспособления).
4. Биосферно-биогеоценотический (элементарными структурами служат биогеоценозы, элементарными явлениями — смена биогеоценозов).
5. Цитология — наука о клетке. Предмет цитологии, — клетки одноклеточных, а также многоклеточных организмов. Предпосылкой открытия клетки были изобретение микроскопа и использование его для исследования биологических объектов.
1590 г. Я н с е н изобрел микроскоп, в котором большое увеличение обеспечивалось соединением двух линз.
1666 г. Р. Г у к, пользуясь усовершенствованным микроскопом, изучал строение пробки и впервые употребил термин клетка для описания структурных единиц, из которых состоит эта ткань. Он считал клетки пустыми, а живое вещество — это клеточные стенки.
1650-1700 гг. Антони ван Левенгук при помощи простых хорошо отшлифованных линз (200 х) наблюдал «зародыши» и различные одноклеточные организмы, в том числе бактерии. Впервые бактерии были описаны в 1676 г.
1700-1800 гг. Опубликовано много новых описаний и рисунков различных тканей, по преимуществу растительных.
1827 г.Долланд резко улучшил качество линз. После этого интерес к микроскопии быстро возрос и распространился.
1831 г. Р. Б р о у н описал ядро в растительных клетках.
1838— 1839 гг. Ботаник Шлейден и зоолог Ш в а н н объединили идеи разных ученых и сформулировали «клеточную теорию», которая постулировала, что основной единицей структуры и функции в живых организмах является клетка.
1840 г. Пуркинье предложил название протоплазма для клеточного содержимого, убедившись в том, что именно содержимое (а не клеточные стенки) представляют собой живое вещество. Позднее был введен термин цитоплазма (цитоплазма + ядро = = протоплазма).
1855 г. В и р х о в показал, что все клетки образуются из других клеток путем клеточного деления.
1866 г. Геккель установил, что хранение и передачу наследственных признаков осуществляет ядро.
1866—1888 гг. Подробно изучено клеточное строение и описаны хромосомы.
1880— 1883 гг. Открыты пластиды, в частности хлоропласты.
1890 г. Открыты митохондрии.
1898 г. Открыт аппарат Гольджи.
1887— 1900 гг. Усовершенствованы микроскоп, а также методы фиксации, окрашивания препаратов и приготовления срезов.
1900 г. Вновь открыты законы Менделя, забытые с 1865 г., и это дало толчок развитию цитогенетики. Световой микроскоп достиг теоретического предела разрешения.
1930-е гг. Появился электронный микроскоп.
С 1946 г. и по настоящее время электронный микроскоп получил широкое распространение в биологии.
6. Методы цитологии
• Микроскопический метод
• Метод электронной микроскопии
• Методы гистохимии, основанные на избирательном действии реактивов и красителей на вещества цитоплазмы
• Метод дифференциального центрифугирования, который позволяет исследовать химический состав органоидов клетки после их разделения с помощью центрифуги
• Метод рентгеноструктурного анализа дает возможность определить пространственное расположение, физические свойства молекул
• Метод авторадиографии основан на введении в клетку меченых радиоактивных атомов
• Метод замедленной киносъемки (наблюдают деление клетки)
• Генная и клеточная инженерия
Немецкий зоолог Т. Ш в а н н (1810-1882) в 1839 г. опубликовал труд «Микроскопические исследования о соответствии в структуре и росте животных и растений». В этой классической работе были заложены основы клеточной теории. Шванн выдвинул основные положения клеточной теории:
1) клетка является главной структурной единицей всех организмов (растительных и животных);
2) процесс образования клеток обусловливает рост, развитие и дифференцировку растительных и животных тканей.
В 1858 г. вышел в свет основной труд немецкого патолога Р. В и р хо в а (1821-1902) «Целлюлярная патология», в котором он выдвинул положение «каждая клетка от клетки».
7. Положения клеточной теории
1. Все растительные и животные организмы состоят из клеток.
2. Клетки растений и животных гомологичны по своему строению.
3. Каждая клетка от клетки. Новые клетки появляются путем деления исходной материнской клетки.
4. Клетки в многоклеточном организме теряют свою самостоятельность. Они подчиняются единой нейрогуморальной регуляции.
Литература
1. Р.Г.Заяц, И.В. Рачковская и др. Биология для абитуриентов. Минск, «Юнипресс», 2009г., с. 3-7.
2. Л.Н. Песецкая. Биология. Минск, «Аверсэв», 2007г., с.4-14
3. Н.Д. Лисов, Н.А. Лемеза и др. Биология. Минск, «Аверсэв», 2009г, с.4-12
4. Е.И. Шепелевич, В.М. Глушко, Т.В. Максимова. Биология для школьников и абитуриентов. Минск, «УниверсалПресс», 2007г., с.5-7, 58-60.
Лекция 2. Структурная организация клетки
Вопросы:
1. Структурные компоненты клетки.
2. Наружная клеточная мембрана, ее строение и роль в обмене веществ.
3. Цитоплазма и органоиды.
4. Ядро, хромосомы. Понятие о кариотипе и генотипе.
1. Типичная клетка состоит из плазматической мембраны, цитоплазмы с различными органеллами и ядра. Растительные клетки, кроме того, имеют и вакуоли, хорошо оформленную клеточную стенку и пластиды.
2. По другой классификации ядро относят к самым крупным органеллам клетки.
Клеточные мембраны играют важную роль:
1) отделяют клеточное содержимое от внешней среды;
2) регулируют обмен между клеткой и средой и делят клетки на отсеки;
3) некоторые химические реакции протекают на мембранах (световые реакции фотосинтеза в мембранах гран хлоропластов или окислительное фосфорилирование в митохондриях);
4) здесь же располагаются и рецепторные участки для распознавания внешних стимулов, поступающих из окружающей среды.
С конца прошлого века известно, что клеточные мембраны ведут себя не так, как полупроницаемые мембраны. Клеточные мембраны обладают избирательной проницаемостью — одни вещества пропускают, а другие не пропускают.
Ранние работы по проницаемости мембран показали, что органические растворители — спирт, эфир или хлороформ — проникают сквозь мембрану быстрее, чем вода. Это говорит о том, что в мембранах есть неполярная часть — липиды. Позже выяснилось: мембраны состоят из белков и липидов.
Липиды в мембранах представлены фосфолипидами, гликолипидами и стеролами. У фосфолипидов (соединений, содержащих фосфатную группу) молекулы состоят из полярной и неполярной части — полярной головы и двух неполярных хвостов. Гликолипиды — продукт соединения липидов с углеводами. Они тоже состоят из полярной головы и неполярных хвостов. Стеролами называют спирты, относящиеся к классу стероидов. Наиболее распространен среди них холестерол. Его молекулы полностью неполярны и в этом его отличие от фосфолипидов и гликолипидов.
Давсон и Даниелли в 1935 г. высказали предположение, что в клеточных мембранах имеется липидный слой, заключенный между двумя слоями белка. В 1959 г. Робертсон, объединив имеющиеся в то время данные, выдвинул гипотезу о строении «элементарной мембраны»:
а) все мембраны имеют толщину около 7,5 нм;
б) в электронном микроскопе они представляются трехслойными;
в) центральный липидный бислой заключен между двумя слоями белка.
В настоящее время наибольшее признание получила жидкостно-мозаичная модель организации мембраны, предложенная Зингером и Николсом в 1972 г., согласно которой белковые молекулы, плавающие в жидком липидном бислое, образуют в нем как бы своеобразную мозаику (рис. 2).
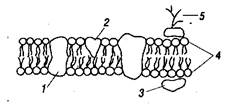
Рис. 2. Схема жидкостно-мозаичной модели мембраны
Белки представлены: интегральными белками /, которые пронизывают всю толщу мембраны; полуинтегральными белками 2, молекулы которых наполовину погружены в мембрану, выступая с внешней или внутренней ее стороны; периферическими белками 3, которые располагаются на поверхности билипидного слоя и связаны с «головками» липидных молекул 4.
Предполагается, что в белковых молекулах или между соседними белковыми молекулами имеются гидрофильные каналы, или поры. Эти поры пронизывают мембрану, так что по ним сквозь мембрану могут проходить полярные молекулы, которые без таких пор пройти бы не могли — липидный компонент мембраны не пропустил бы их в клетку. Некоторые белки действуют как переносчики. В мембранах содержатся ферментные белки, специфические рецепторы, переносчики электронов, преобразователи энергии, участвующие в фотосинтезе и дыхании.
Кроме того, в мембранах имеются гликопротеины. У них на свободных поверхностях находятся гликозильные группы — разветвленные олигосахаридные цепи 5, напоминающие антенны. Эти «антенны» состоят из нескольких моносахаридных остатков. Функция «антенн» связана с распознаванием внешних сигналов, которое важно для клеток по многим причинам. Распознающие участки двух соседних клеток могут связываться друг с другом, обеспечивая сцепление клеток. Благодаря этому клетки правильно ориентируются и образуют ткани в процессе дифференцировки. С распознаванием связана и деятельность различных регулятор-ных систем, а также иммунный ответ, в котором гликопротеины играют роль антигенов. Распознающие участки имеются и у некоторых молекул, находящихся в растворе, благодаря чему они избирательно поглощаются клетками с комплементарными распознающими участками. Присоединение углеводных участков к белкам осуществляется в аппарате Гольджи. Сахара, таким образом, могут функционировать как информационные молекулы, т. е. в этом смысле их можно сравнивать с белками и нуклеиновыми кислотами.
Жизнедеятельность клетки как открытой системы зависит от проникновения внутрь необходимых ей веществ и выделения продуктов обмена из клетки. Вещества могут поступать в клетку через мембрану следующими путями:
I — пассивная диффузия — по градиенту концентрации (молекулы воды и другие небольшие молекулы);
II — облегченная диффузия, когда транспорту способствует белок-переносчик (сахар, аминокислоты, нуклеотиды, гормоны);
III — активный транспорт — перенос против градиента концентрации через специальные каналы, для чего необходима энергия АТФ (натрий—калиевый насос);
IV — эндоцитоз (фаго- и пиноцитоз) — поступление крупных молекул и воды вместе с участками плазмолеммы, образующими эндосому.
Сравнительно недавно выяснилось, что у большей части клеток в плазматической мембране действует натриевый насос активно выкачивающий натрий из клетки. Обычно, хотя и не всегда, натриевый насос сопряжен с калиевым насосом, активно поглощающим ионы калия из внешней среды и переносящим их в клетку. Такой объединенный насос называют натрий-калиевым насосом (Na\ К-насос). О его физиологическом значении свидетельствует тот факт, что более трети АТФ, потребляемого животной клеткой в состоянии покоя, расходуется на перекачивание натрия и калия. Это необходимо для сохранения клеточного объема (осмо-регуляция), для поддержания электрической активности в нервных и мышечных клетках и для активного транспорта некоторых других веществ (сахаров, аминокислот). Высокие концентрации калия требуются также для белкового синтеза, гликолиза, фотосинтеза и для некоторых других жизненно важных процессов. Насос— это особый белок, локализующийся в мембране таким образом, что он пронизывает всю ее толщу. С внутренней стороны мембраны к нему поступают натрий и АТФ, а с наружной — калий. На каждые два поглощенных иона калия из клетки выводится три иона натрия. Выкачиваемый из клетки натрий обычно пассивно диффундирует в клетку. Однако мембрана мало проницаема для натрия. Для ионов калия мембраны приблизительно в 100 раз более проницаемы.
3. Цитоплазма состоит из:
• жидкого содержимого — гиалоплазмы;
• органоидов;
• включений.
Гиалоплазма — гетерогенный коллоидный раствор, обеспечивающий взаимосвязь всех органоидов клетки и процессы ее жизнедеятельности. Гиалоплазма обеспечивает клетке вязкость, эластичность, сократимость, внутреннее движение. В ней расположены микротрубочки — особые белковые структуры, выполняющие опорную функцию клетки;
Органоиды — специализированные постоянные компоненты цитоплазмы, которые обладают определенным строением и выполняют ту или иную функцию жизнедеятельности клетки. Делятся на две группы:
• органоиды общего назначения (митохондрии, комплекс Гольджи, пластиды, клеточный центр, вакуоли, эндоплазматическая сеть (ЭС), рибосомы, лизосомы);
• органоиды специального назначения (реснички, жгутики, нейрофибриллы, миофибриллы).
Цитоплазматические включения — это непостоянные структуры в цитоплазме, представляющие собой продукты жизнедеятельности клеток. По своему биологическому значению включения могут быть условно разделены на три основные группы:
а) трофические — запасные питательные вещества (крахмал и белковые зерна, капли жира в растительных клетках; гликоген и жир — в животных клетках);
б) секреторные — продукты жизнедеятельности железистых клеток (гормоны, ферменты, секреты);
в) экскреторные — конечные продукты обмена растительных и животных клеток (кристаллы оксалата кальция, гипса, мочевой кислоты).
Органоиды немембранного строения — рибосомы и клеточный центр. Рибосомы — ультрамикроскопические органеллы, состоящие из двух частей — субъединиц, состоящих из белка и рРНК. Субъединицы образуются в ядрышке. Это универсальные органеллы всех клеток животных и растений. Находятся в цитоплазме и на мембранах ЭС. В рибосомах синтезируются белки.
Клеточный центр состоит из двух центриолей. Каждая имеет цилиндрическую форму, стенки образованы девятью триплетами микротрубочек, а в середине находится однородное вещество. Центриоли расположены перпендикулярно друг другу. Клеточный центр принимает участие в делении клеток животных и низших растений.
Органоиды, имеющие двухмембранное строение, — это митохондрии и пластиды. У митохондрий внешняя мембрана гладкая, внутренняя образует различной формы выросты — кристы. В матриксе митохондрий (полужидком веществе) находятся ферменты, рибосомы, ДНК, РНК. Это — дыхательный и энергетический центр, где на кристах идет синтез АТФ.
Пластиды (лейкопласты, хлоропласта, хромопласта) — это органоиды различной окраски и функций, характерные только для растений. Лейкопласты — бесцветны, служат местом отложения запасных питательных веществ. Хлоропласта имеют зеленую окраску благодаря хлорофиллу, который располагается в мембранах тилакоидов гран. Здесь осуществляется фотосинтез, т. е. эти органеллы способны из С02 и Н20 при наличии световой энергии создавать органические вещества — углеводы и свободный кислород. Хромопласты имеют красную, оранжевую, желтую окраску. Придают окраску цветам, осенним листьям, плодам.
Органоиды одномембранного строения: эндоплазматическая сеть, аппарат Гольджи, лизосомы. ЭС — ультрамикроскопическая система мембран, образующих трубочки, канальцы, цистерны, пузырьки. Гранулярная ЭС несет рибосомы, гладкая — лишена их. Обеспечивает транспорт веществ, гранулярная ЭС несет рибосомы и участвует в синтезе белка. В каналах ЭС образуются сложные молекулы белка, синтезируются жиры, транспортируется АТФ.
Аппарат Гольджи обнаружен в 1898 г., состоит из стопочки плоских цистерн, по краям которых ответвляются трубочки, отделяющие мелкие пузырьки. В цистернах накапливаются продукты синтеза, распада и вещества, поступившие в клетку, а также вещества, которые выводятся из клетки. В растительной клетке пузырьки Гольджи участвуют в построении клеточной стенки.
Лизосомы — органоиды округлой формы. В них находятся лизирующие (растворяющие) ферменты, синтезированные на рибосомах. Они переваривают пищу, попавшую в животную клетку, в клетках любых организмов осуществляют автолиз (саморастворение органелл). Например, у лягушонка рассасывается хвост.
4. Ядро, хромосомы. Понятие о кариотипе
Составными частями ядра эукариотов являются:
• ядерная оболочка — двухслойная пористая оболочка (кариолемма);
• ядерный сок (кариоплазма) содержит ферменты, необходимые для синтеза нуклеиновых кислот и рибосом;
• ядрышки — не постоянные структуры, они исчезают в начале деления клетки и появляются к концу его. Ядрышки содержат белки и РНК;
• хромосомы — наиболее важные структурные компоненты ядра.
Строение хромосом. Хромосомы могут находиться в двух структурно-функциональных состояниях: в конденсированном (спирализованном) и деконденсированном (деспирализованном). В неделящейся клетке хромосомы не видны, обнаруживаются лишь глыбки и гранулы хроматина. Ко времени деления клетки происходит конденсация (спирализация) хроматина и при митозе хромосомы хорошо видны. ,
Хроматин представляет собой комплекс ДНК и белков. В состав хроматина входит два типа белков: гистоновые (белки основной природы) и нееистоновые (белки кислые).
Уровни упаковки генетического материала:
• нуклеосомный;
• супернуклеосомный;
• хроматидный;
• уровень метафазной хромосомы.
Нуклеосома (глобула) состоит из восьми молекул белков, вокруг которой двойная нить ДНК делает около двух витков. Здесь длина молекулы ДНК уменьшается в 5 раз. Супернуклеосомный уровень. Нуклеосомная нить конденсируется, присоединяя белки, и образует спираль. Она обнаруживается как в интерфазных, так и в митотических хромосомах. Этот уровень дает укорочение нити еще в 6 раз. Хроматидный. Супернуклеосомная нить способна образовывать петли и изгибы. Она составляет основу хроматиды. Уровень дает укорочение нити в 10—20 раз. Уровень метафазной хромосомы. Хроматиды способны спирализоваться с образованием эухроматиновых (слабо спирализованных) и гетерохроматиновых (сильно спирализованных) участков. Укорочение в 10 ООО раз.
Хромосомы были детально изучены В. Вальдейером в 1888 г. За способность интенсивно окрашиваться они получили название хромосом. Хромосома состоит из двух хроматид и после деления ядра становится однохроматидной. К началу следующего деления у каждой хромосомы достраивается вторая хроматида. Форма хромосомы зависит от положения первичной перетяжки — центромеры. Это механический центр хромосомы, к которому прикрепляются нити веретена деления. Первичная перетяжка делит хромосому на два плеча. В зависимости от места расположения центромеры различают три типа хромосом:
• метацентрические хромосомы имеют плечи равной величины;
• субметацентрические — имеют плечи неравной длины;
• акроцентрические имеют палочковидную форму с очень коротким, почти незаметным вторым плечом.
Могут возникнуть и телоцентрические хромосомы в результате отрыва одного плеча. Центромера у них находится на конце хромосомы. В нормальном кариотипе такие хромосомы не встречаются.
Классификация хромосом была предложена в 1969 г. на международной конференции в Денвере.
Различают гаплоидный — одинарный набор хромосом (в зрелых половых клетках) и диплоидный — двойной (в соматических клетках). Совокупность.генов в гаплоидном наборе хромосом называется геномом. Диплоидный набор хромосом клетки, характеризующийся их числом, величиной и формой, называется кариотипом.
Правила хромосом
1. Правило индивидуальности. Гомологичные хромосомы одинаковы по форме и величине. Каждая пара хромосом характеризуется своими особенностями. В этом выражается правило индивидуальности.
2. Правило парности. У организмов, как правило, число хромосом четное. Это связано с тем, что хромосомы составляют пары. У лошадиной аскариды одна пара хромосом, у дрозофилы — четыре, у человека — 23.
3. Правило постоянства числа хромосом (примеры в п. 2).
4. Правило непрерывности хромосом, т.е. каждая хромосома воспроизводит себе подобную, что обеспечивается редупликацией и делением клетки.
Сходство и отличие растительной и животной клетки
Общие признаки
1. Единство структурных систем- — цитоплазмы и ядра.
2. Единство процессов обмена веществ и энергии.
3. Единство принципа наследственного кода.
4. Универсальное мембранное строение.
5. Единство химического состава.
6. Сходство процесса деления клеток.
Отличительные признаки
| Для сравнения | Растительная клетка | Животная клетка |
| Пластиды | Хлоропласты, хромопласты, лейкопласты | Отсутствуют |
| Способ питания | Автотрофный (фототроф- ный, хемотрофный) | Гетеротрофный (сапрофит- ный, паразитический) |
| Синтез АТФ | В хлоропластах, митохондриях | В митохондриях |
| Центриоли | У низших растений | Во всех клетках |
| Целлюлозная клеточная оболочка | Расположена-снаружи от клеточной мембраны | Отсутствует |
| Вакуоли | Крупные полости, запол- ненные клеточным соком | Сократительные, пищева- рительные, выделительные вакуоли. Обычно мелкие |
| Разделение при митозе | Разделение начинается во внутренней области мате- ринской клетки, здесь мел- кие пузырьки ЭПС слива- ются, образуя клеточную мембрану | В экваториальной зоне об- разуется перетяжка, кото- рая все больше углубля- ясь, отделяет сестринские клетки друг от друга |
Тест 2
1. В мембранах эукариот липидов:
а) один слой;
б) два слоя;
в) три слоя.
2. Рецепторная функция мембраны связана с:
а) белками илипидами;
б) липидами и углеводами;
в) белками и углеводами.
3. Липиды по отношению к воде являются соединениями:
а) гидрофильными;
б) гидрофобными.
4. Выведение веществ из клетки называется:
а) экзоцитозом;
б) фагоцитозом;
в) эндоцитозом;
г) пиноцитозом.
5. Ядро представляет собой:
а) двухмембранную структуру;
б) одномембранную структуру;
в) немембранную структуру.
6. Строение рибосомы:
а) одномембранное;
б) двухмембранное;
в) немембранное.
7. Хроматин — это:
а) молекула ДНК;
б) молекула РНК;
в) комплекс ДНК-белок;
г) только белок, способный связываться с ДНК.
8. Центромеры в хромосомах располагаются:
а) только посередине;
б) только по периферии;
в) в любом месте хромосомы.
9. Гомологичные хромосомы одинаковы по форме и величине; Каждая пара хромосом характеризуется своими особенна»' стями. В этом выражается правило:
а) индивидуальности;
б) парности;
в) постоянства числа хромосом;
г) непрерывности.
10. Разделение цитоплазмы начинается во внутренней области материнской клетки, здесь мелкие пузырьки эндоплазматической сети сливаются, образуя клеточную мембрану. Это — отличительные признаки митоза:
а) растительной клетки;
б) животной клетки.
Литература
1. Р.Г.Заяц, И.В. Рачковская и др. Биология для абитуриентов. Минск, «Юнипресс», 2009г., с. 7-12,534-554.
2. Л.Н. Песецкая. Биология. Минск, «Аверсэв», 2007г., с.5-23.
3. Н.Д. Лисов, Н.А. Лемеза и др. Биология. Минск, «Аверсэв», 2009г, с.25-29.
4. Е.И. Шепелевич, В.М. Глушко, Т.В. Максимова. Биология для школьников и абитуриентов. Минск, «УниверсалПресс», 2007г., с.7-9, 20-36.
Лекция 3. Обмен веществ и превращение энергии в клетке.
Вопросы:
1. Энергетический обмен.
2. Биосинтез белков.
3. Фотосинтез.
1. Совокупность реакций ассимиляции и диссимиляции, лежащих в основе жизнедеятельности и обусловливающих связь организма с окружающей средой, называется обменом веществ.
Энергетическим обменом, или диссимиляцией, или катаболизмом называется совокупность реакций ферментативного расщепления (белков, жиров, углеводов) и образования соединений, богатых энергией (АТФ).
АТФ — мононуклеотид, состоящий из азотистого основания аденина, пятиуглеродного моносахарида рибозы и трех остатков фосфорной кислоты, которые соединены друг с другом макроэргическими связями. Отщепление одной молекулы фосфорной кислоты сопровождается выделением 40 кДж энергии.
Выделяют три этапа энергетического обмена:
• подготовительный;
• бескислородный;
• кислородный.
Первый этап — подготовительный, называемый также пищеварением, осуществляется в пищеварительном тракте животных и человека или в цитоплазме клеток всех живых существ. На этом этапе крупные молекулы полимеров расщепляются на мономеры: белки — на аминокислоты, полисахариды — на простые сахара (моносахариды), жиры — на жирные кислоты и глицерин, нуклеиновые кислоты — на нуклеотиды. При этом выделяется небольшое количество энергии, которая рассеивается в виде теплоты.
Бескислородный (гликолиз) этап протекает в цитоплазме клеток. Мономеры, образовавшиеся на первом этапе, подвергаются дальнейшему расщеплению без участия кислорода. Например, при гликолизе (расщеплении глюкозы, происходящем в животных клетках) одна молекула глюкозы расщепляется на две молекулы пировиноградной кислоты (С3Н403), которая в мышечных клетках восстанавливается до молочной кислоты. При этом выделяется около 200 кДж энергии. Из них 80 кДж идет на синтез двух молекул АТФ, а остальная энергия (около 120 кДж) рассеивается в виде тепла. Суммарное уравнение этой реакции:
С6Н1206 + 2АДФ + 2Н3Р04 ═ 2С3Н603 + 2АТФ + 2Н20.
В клетках растительных организмов и некоторых дрожжевых грибков распад глюкозы идет путем спиртового брожения (до этилового спирта). У анаэробных организмов (некоторые бактерии, внутрикишечные паразиты) этот этап является конечным.
Кислородный (аэробный) этап имеет место только у аэробных организмов. Он заключается в дальнейшем окислении молочной (или пировиноградной) кислоты до конечных продуктов — углекислого газа и воды.
Уравнение кислородного этапа имеет вид:
2С3Н603 + 602 + 36Н3Р04 + 36АДФ ═ 36АТФ + 6С02 + 42Н20.
Этот процесс протекает в митохондриях с участием ферментов и кислорода. На первых стадиях кислородного этапа от молочной кислоты постепенно отщепляются протоны и электроны и накапливаются по разные стороны внутренней мембраны митохондрии, создавая разность потенциалов.
Когда разность потенциалов достигает критического значения, протоны, проходя по каналам АТФ-синтетаз, отдают свою энергию для синтеза АТФ. На этом этапе синтезируется 36 молекул АТФ.
Суммарное уравнение анаэробного и аэробного этапов:
С6Н,206 + 38АДФ + 38Н3Р04 + 602 ═ 38АТФ + 6С02 + 44Н20;
С6Н1206 + 602 ═6С02 + 6Н20 + 38АТФ.
Таким образом, в ходе второго и третьего этапов энергетиче ского обмена при расщеплении одной молекулы глюкозы образуется 38 молекул АТФ. Медиков особенно интересуют вещества, повышающие проницаемость мембран, как бы перфорирующие их. Если мембрана перфорирована, то часть ионов Н+ ускользает из протонного резервуара, минуя канал, где АТФ-синтетаза могла бы использовать энергию для синтеза АТФ. Перфорацию митохондриальных мембран вызывают, например, динитрофенолы — вещества желтого цвета, которые одно время пытались добавлять к различным мучным изделиям, чтобы придать им более «сдобный» вид. Предпринимались так же попытки применить эти вещества в качестве средства от ожирения, но попытки эти окончились трагически — несколько человек от такого «лечения» умерло, так как их организм перестал вырабатывать достаточное количество АТФ. Подобное разобщение процесса переноса электронов и синтеза АТФ иногда имеет место у животных и в норме. У животных, впадаю щих в зимнюю спячку, в клетках бурой жировой ткани большая часть энергии, высвобождаемой во время переноса электронов, расходуется не на синтез АТФ, а на производство тепла.
Пластический обмен, ассимиляция, — это совокупность реакций синтеза, направленных на образование структурных частей клеток и тканей. К пластическому обмену относится биосинтез белков, фотосинтез, синтез нуклеиновых кислот, жиров и углеводов.
2. Биосинтез белка включает два основных процесса: транскрипции и трансляции. Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном. Таким образом, информация о структуре белка находится в ДНК-Процесс списывания информации с ДНК на структуру иРНК, осуществляемый РНК-полимеразой, называется транскрипцией. Информационная РНК является копией не всей молекулы ДНК, а только части ее — одного гена или группы рядом лежащих генов. У прокариот такая группа генов называется опероном (рис. 3).
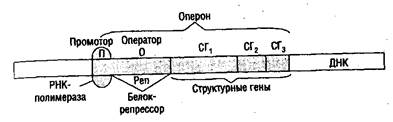
Рис. 3. Схема строения оперона прокариот
В начале каждого оперона находится своего рода посадочная -1 площадка для РНК-полимеразы, называемая промотором. Только присоединившись к промотору РНК-полимераза способна начать синтез иРНК. Следующий участок — оператор, с него начинается операция — синтез иРНК. С оператором взаимодействует специальный белок — репрессор (подавитель). Пока репрессор находится на операторе, РНК-полимераза не может сдвинуться с места и начать синтез иРНК.
В структурных генах закодирована структура ферментов Ф1, Ф2, ФЗ, необходимых, например, для расщепления субстрата (сахара в клетках бактерий).
Дойдя до конца оперона, РНК-полимераза встречает сигнал (в виде определенной последовательности нуклеотидов), означающий конец считывания. Готовая иРНК отходит от ДНК и направляется к месту синтеза белков. В процессе транскрипции можно выделить четыре стадии:
1) связывание РНК-полимеразы с промотором;
2) инициация — начало синтеза. Она заключается в образовании первой фосфодиэфирной связи между АТФ и вторым нуклеотидом синтезирующейся молекулы иРНК;
3) элонгация — рост цепи РНК, т. е; последовательное присоединение нуклеотидов друг к другу. Скорость элонгации достигает 50 нуклеотидов в секунду;
4) терминация — завершение синтеза иРНК.
Благодаря процессу транскрипции в клетке осуществляется передача информации от ДНК к белку:
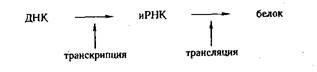
Перевод информации с «языка» нуклеотидов на «язык» аминокислот осуществляется с помощью генетического кода. Код — это система символов для перевода одной формы информации в другую.
Свойства кода
• Код триплетен. Это означает, что каждая из 20 аминокислот зашифрована последовательностью трех нуклеотидов, называемых триплетом или кодоном.
• Код вырожден. Это означает, что каждая аминокислота шифруется более чем одним кодоном.
• Код однозначен. Каждый кодон шифрует только одну аминокислоту.
• Между генами имеются знаки «препинания» — кодоны-терминаторы. Код является неперекрывающимся, т. е. один и тот же нуклеотид не может входить одновременно в состав двух соседних триплетов.
• Внутри гена нет «знаков препинания», поскольку генетический код подобен языку.
• Код универсален. Генетический код един для всех живущих на Земле существ.
Трансляция
Синтез полипептидных цепей белков по матрице иРНК, выполняемый рибосомами, называется трансляцией. Аминокислоты, из которых синтезируются белки, доставляются к рибосомам с помощью специальных тРНК. Это небольшие молекулы, состоящие из 70-90 нуклеотидов, напоминающие по форме клеверный лист. На вершине «листа» каждой тРНК имеется последовательность трех нуклеотидов, комплементарных нуклеотидам кодона в иРНК, их называют антикодоном. Фермент кодаза опознает тРНКи присоединяет к «черешку листа» аминокислоту, комплементарную антикодону. На образование этой связи затрачивается энергия одной молекулы АТФ.
Внутри рибосомы в каждый данный момент находится всего два триплета иРНК. Рибосома движется относительно иРНК только в одном направлении, перемещаясь на один триплет. Синтез белковой молекулы происходит в большой субъединице, где против одного триплета расположен аминоацильный центр (акцепторный участок), а против другого — пептидильный (донорный) участок, где формируются пептидные связи.
Когда на рибосоме оказывается один из трех триплетов (УАА, УАГ, УГА), являющихся «знаками препинания» между генами, ни одна тРНК не может занять место в акцепторном участке, так как не существует антикодонов, комплементарных нуклеотидам «знаков препинания». Оторвавшейся в донорном участке полипептидной цепи не к чему присоединиться в акцепторном участке и она покидает рибосому. Синтез белка завершен. А начинается синтез белка с того, что с кодоном АУГ, расположенным на первом месте в копии каждого гена, взаимодействует антикодон особой тРНК, соединенной с формилметионином. Эта аминокислота выполняет роль заглавной буквы в предложении, с нее в бактериальной клетке начинается синтез любой далипептидной цепи. Когда триплет АУГ стоит внутри копии гена, он кодирует аминокислоту метионин.
Информационная РНК часто одновременно проходит не по одной, а по нескольким рибосомам. Такую структуру, объединенную одной молекулой иРНК, называют полисомой. В 1961 г. французские ученые Ф. Жако б, А. Львов и Ж. Моно опубликовали результаты своих исследований по регуляции белкового синтеза у бактерий. За эту работу они были удостоены Нобелевской премии.
3. Фотосинтез
По типу ассимиляции все клетки делят на две группы — автотрофные и гетеротрофные. Автотрофные клетки способны к самостоятельному синтезу органических соединений за счет углекислого газа, воды и энергии света или энергии химических связей (зеленые растения и некоторые бактерии). Гетеротрофные клетки не могут синтезировать органические вещества из неорганических, они их получают извне в виде пищи (животные, грибы, большая часть бактерий, некоторые высшие растения — подъельник, заразиха, омела).
Фотосинтез — процесс образования органических соединений из диоксида углерода и воды с использованием энергии света. Свет улавливается хлорофиллом-а, встроенным во внутреннюю мембрану пластид у эукариот или в складки цитоплазматической мембраны прокариот.
Фотосинтез подразделяется на световую и темновую фазы. Световая фаза — это этап, на котором энергия света, поглощенная хлорофиллом, преобразуется в электрохимическую энергию в цепи переноса электронов. Осуществляется на свету, в мембранах гран при участии белков-переносчиков и АТФ-синтетазы.
Реакции световой фазы:
• Возбуждение электронов хлорофилла квантами света и переход их на более высокий уровень.
• Фотолиз воды, происходящий при участии квантов света:
2Н20 ═ 4Н* + 4е + 02.
• Протоны накапливаются в Н+-резервуаре внутри граны. Их накопление на внутренней стороне мембраны приводит к нарастанию разности потенциалов.
• При достижении критической разности потенциалов протоны движутся по каналу в АТФ-синтетазе из тилакоида в строму. На выходе из протонного канала создается высокий уровень энергии, которая идет на синтез АТФ:
АДФ + Ф ═ АТФ.
•. Восстановление акцепторов электронов — НАДФ* до НАДФН2:
2Н+ + 4е + НАДФ+ ═ НАДФН2.
Результаты световой фазы:
1) образование кислорода при фотолизе воды;
2) синтез АТФ;
3) восстановление НАДФ.
Темновая фаза — процесс преобразования углекислого газа в глюкозу в строме хлоропластов с использованием энергии АТФ и НАДФН2.
Реакции темновой фазы — последовательные преобразования углекислого газа в углеводы:
• Фиксация молекулы С02 ] ,5-рибулозодифосфатом при участии ферментов.
• Постепенное восстановление диоксида до глюкозы при участии АТФ и НАДФН2 (цикл Кальвина).
• Образование, кроме молекул глюкозы, аминокислот, нуклеотидов, спиртов.
Суммарное уравнение фотосинтеза
6С02 + 6Н20 ═ С6Н|1206 + 602.
Значение фотосинтеза:
• в процессе фотосинтеза образуется свободный кислород, необходимый для дыхания организмов;
• фотосинтез обеспечивает производство исходных органических веществ, а следовательно, пищу для всех живых существ;
• фотосинтез способствует снижению концентрации диоксида углерода в атмосфере.
В изучение процесса фотосинтеза большой вклад внесли К. А. Т и м и р я з е в, американский ученый М. К а л ь в и н, австралийские М. Д. X е т ч и К. Р. С л э к, а также наши белорусские ученые Т. Н. Год н е в и А. А. Ш л ы к (Т. Н. Годнев — физиолог растений. Его работы посвящены биохимии фотосинтезирующих пигментов. Т. Н. Годневу принадлежит теория образования хлорофилла.)
Тест 3
1. Ассимиляция называется пластическим обменом потому:
а) что создаются органические вещества;
б) что расщепляются органические вещества.
2. Синтез АТФ происходит:
а) на кристах;
б) в матриксе;
в) на наружной мембране митохондрии.
3. Синтез АТФ в хлоропластах связан:
а) со световой фазой фотосинтеза;
б) с темновой фазой фотосинтеза;
в) с синтезом белков.
4. Свободный кислород образуется в ... стадии фотосинтеза.
а) темновой;
б) световой;
в) постоянно.
5. В каких структурах растительной клетки накапливается крахмал:
а) в митохондриях;
б) в хлоропластах;
в) в лейкопластах?
6. Какая из структур ядра содержит информацию о синтезе одного белка:
а) молекула ДНК;
б) ген;
в) нуклеотид;'
г) триплет нуклеотидов
7. Короткая последовательность
ДНК 5'-AGGATGCTA-3' может полностью гибридизоваться с:
а) 5'-AGGATGCTA-3';
б) 5'-UGGUACGAU-3';
в) 5'-ATGGTAGGA-3';
г) 5'-TAGCATCCT-3'.
8. Какая из ядерных структур принимает участие в сборке субъединиц рибосом?
а) ядерная оболочка;
б) ядрышко;
в) ядерный сок.
9. Генетическая информация определяется последовательностью:
а) фосфатных групп;
б) сахарных групп;
в) нуклеотидов;
г) аминокислот.
10. Геном эукариот отличается:
а) высокой плотностью кодирования;
б) наличием последовательностей, которые не кодируют никаких известных функций;
в) наличием только уникальных генов;
г) наличием только повторяющихся последовательностей.
Литература
1. Р.Г.Заяц, И.В. Рачковская и др. Биология для абитуриентов. Минск, «Юнипресс», 2009г., с. 578-597.
2. Л.Н. Песецкая. Биология. Минск, «Аверсэв», 2007г., с.23-35.
3. Н.Д. Лисов, Н.А. Лемеза и др. Биология. Минск, «Аверсэв», 2009г, с.33-37.
4. Е.И. Шепелевич, В.М. Глушко, Т.В. Максимова. Биология для школьников и абитуриентов. Минск, «УниверсалПресс», 2007г., с.37-50.
Лекция 4. Размножение и индивидуальное развитие организмов.
Вопросы:
1. Деление клеток. Митоз. Амитоз. Мейоз.
2. Сравнение митоза и мейоза.
3. Размножение.
4. Индивидуальное развитие.
1. Совокупность процессов, протекающих в клетке от момента ее появления до гибели или деления на две дочерние, включая само деление, называется жизненным циклом клетки. Совокупность процессов, протекающих в клетке от одного деления до другого, включая само деление, называется митотическим циклом.
Период в жизнедеятельности клетки от ее образования до начала следующего деления называют интерфазой. В интерфазе различают три периода:
• пресинтетический;
• синтетический;
• постсинтетический.
Пресинтетический период — синтез белка; на деспирализованных молекулах ДНК синтезируется РНК. Синтетический период — синтез ДНК (самоудвоение молекулы ДНК). Построение второй хроматиды, в которую переходит вновь образовавшаяся молекула ДНК: получаются двухроматидные хромосомы. В постсинтетический период происходит синтез белка, накопление энергии, подготовка клетки к делению.
Фазы митоза
1. Профаза. Двухроматидные хромосомы спирализуются, ядрышки растворяются, центриоли расходятся, ядерная оболочка растворяется, образуются нити веретена деления.
2. Метафаза (фаза скопления хромосом). Нити веретена деления присоединяются к центромерам хромосом, двухроматидные хромосомы сосредоточиваются на экваторе клетки.
3. Анафаза (фаза расхождения хромосом). Центромеры делятся, однохроматидные хромосомы растягиваются к полюсам клетки.
4.Телофаза — фаза окончания деления. Однохроматидные хромосомы деспирализуются, сформировывается ядрышко, восстанавливается ядерная оболочка, на экваторе начинает закладываться перегородка между клетками, растворяются нити веретена деления.
Иногда выделяют прометофазу. Эта фаза начинается с разрушения ядерной оболочки.
Биологическое значение митоза заключается в точном идентичном распределении дочерних хромосом с содержащейся в них генетической информацией между ядрами дочерних клеток.
Первое опиеание отдельных фаз митоза сделал И. Д. Чистяков в 1874 г., а подробное описание принадлежит Ф. Ф л е м и н гу в 1878—1882 гг. в животных клетках и Э. С т р а с бургеру (1875, 1880 гг.) в растительных.
Амитоз или прямое деление представляет собой деление ядра без спирализации хромосом и без равномерного их распределения, так как митотический аппарат не образуется. Амитоз описал Р. Р е м а к в 1841 г. Прямое деление характеризуется первоначально перешнуровкой ядрышка, затем ядра и цитоплазмы. Ядро может делиться на две равномерные части (равномерный амитоз) или на две неравномерные части (неравномерный амитоз), либо ядро делится на несколько частей (шизогония у малярийного плазмодия). Иногда после деления ядра цитоплазма не делится и возникают многоядерные клетки (амитоз без цитотомии). В зависимости от факторов, обусловивших амитоз, выделяют три его вида (Жилкин Л. Н., 1966 г):
1. Генеративный амитоз отмечается при делении высокоспециализированных полиплоидных клеток. Наблюдается у инфузории при делении макронуклеуса, в некоторых клетках млекопитающих (печени, эпидермиса).
2. Реактивный амитоз выявляется при различных повреждающих воздействиях (γ-лучи), нарушении обменных процессов (голодании, нарушении нуклеинового обмена). Этот вид амитоза обычно не завершается цитотомией и приводит к образованию многоядерных клеток. Вероятно, его следует рассматривать как компенсаторную реакцию, приводящую к увеличению поверхности обмена между ядром и цитоплазмой.
3. Дегенеративный амитоз возникает в стареющих клетках с угасающими жизненными свойствами. Этот вид представлен фрагментацией и почкованием ядер. Он не имеет отношения к репродукции клеток. Появление дегенеративных форм амитоза служит одним из признаков некробиотических процессов.
Биологическое значение амитоза: увеличение клеточной массы с наименьшей затратой энергии и веществ.

Эндомитоз — один из видов митоза, суть которого заключается в редупликации хромосом без разрушения ядерной оболочки и без деления клетки. Вследствие этого в клетке происходит умножение числа хромосом, иногда в десятки раз по сравнению с исходным. Эндомитоз встречается в интенсивно функционирующих клетках различных тканей: клетки печени, в тканях нематод, насекомых, ракообразных, в корешках некоторых растений. Допускают, что эндомитоз возникает в процессе эволюции, как один из вариантов митоза.
Частный случай эндомитоза — политения. Иногда воспроизведение хромосом происходит без увеличения их числа в клетке. Каждая хромосома многократно удваивается, но дочерние хроматиды остаются связанными между собой. Явление и получило название политении. Оно приводит к увеличению массы ядра и цитоплазмы, соответственно числу удвоений хромосом. Политения встречается у двукрылых насекомых, инфузорий, некоторых растений. В клетках слюнных желез дрозофилы из-за политенизации хромосомплоидность достигает 1024 ядра.
Мейоз — это процесс деления специализированных диплоидных клеток, приводящий к уменьшению числа хромосом вдвое. Мейоз состоит из двух последовательных делений (редукционного и эквационного), которым предшествует однократная репликация ДНК- Интерфаза мейоза аналогична интерфазе митоза.
Редукционное деление. В профазе мейоза I гомологичные хромосомы коньюгируют и обмениваются гомологичными участками (кроссинговер). Профаза I состоит из 5 стадий:
. • Лептотена — реплицированные хромосомы конденсируются.
• Зиготена — начинается коньюгация гомологичных хромосом. Образуются биваленты или тетрады, состоящие их 4-х сестринских хроматид.
• Пахитена — стадия, на которой происходит кроссинговер.
• Диплотена — коньюгировавшие хромосомы разделяются, хромосомы биваленты отодвигаются друг от друга, но продолжают быть связанными хиазмами — местами, где произошел кроссинговер.
• Диакинез — ядерная оболочка и ядрышки исчезают. Каждый бивалент состоит из 4-х хроматид. Сестринские хроматиды соединены центромерой, несестринские — хиазмами.
Далее следуют метафаза I, анафаза I, телофаза I. В телофазе мейоза I каждая дочерняя клетка получает гаплоидный набор хромосом и сразу же наступает мейоз II.
Эквационное деление состоит из профазы II, метафазы II, анафазы II, телофазы II и цитокинеза. Профаза II бывает очень короткой или вообще не наступает. В метафазе II хромосомы, состоящие их двух хроматид, располагаются в экваториальной плоскости, а в анафазе II хроматиды расходятся к противоположным полюсам. В телофазе II образуются четыре гаплоидные клетки.
Биологическое значение мейоза
1. Мейоз обеспечивает поддержание постоянства числа хромосом. Если бы не происходило редукции числа хромосом при га-метогенезе, то из поколения в поколение их число возрастало бы и был бы утрачен один из существенных признаков вида — постоянство числа хромосом.
2. При мейозе образуется большое количество различных новых комбинаций негомологичных хромосом. В диплоидном наборе они двойного происхождения — одна хромосома от отца, а другая от матери.
3. В процессе кроссинговера также происходит рекомбинация генетического материала. Этим достигается еще большая степень перекомбинации наследственного материала.
2. Сравнение митоза и мейоза
Сходство
1. Имеют одинаковые фазы деления.
2. Перед митозом и мейозом происходит самоудвоение хромосом, спирализация и удвоение молекул ДНК.
Отличие
1. В митозе одно деление, в мейозе — два сменяющих друг друга деления.
2. В метафазе митоза по экватору выстраиваются удвоенные хромосомы. В мейозе по экватору выстраиваются пары гомологичных хромосом.
3. В митозе нет коньюгации хромосом. В мейозе гомологичные хромосомы коньюгируют.
4. В митозе между делениями происходит удвоение молекул ДНК (хромосом). В мейозе между первым и вторым делениями нет интерфазы и удвоения молекулы ДНК (хромосом).
5. При митозе образуются две дочерние клетки. При мейозе образуются четыре сперматозоида или одна яйцеклетка.
3. Размножение
Существование особи поддерживается размножением клеток, а существование вида —размножением особей. Размножение необходимо, чтобы поддерживать преемственность поколений. Все размножение можно свести к двум способам: бесполое и половое.
При бесполом размножении участвует одна особь и все потомство является генетически однородным. При половом размножении, как правило, участвуют две особи и полученное потомство несет в себе признаки как одного так и второго организма.
Размножение — одно из основных свойств живого, под которым понимают способность живых организмов воспроизводить себе подобных, обеспечивая непрерывность и преемственность жизни в ряду поколений.
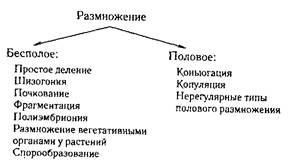
Простое деление характерно для одноклеточных (амебы, жгутиковые, инфузории). Сначала происходит деление ядра, а затем делится цитоплазма. Шизогония или множественное деление встречается у малярийного плазмодия. При шизогонии происходит многократное деление ядра без цитокинеза, а затем и вся цитоплазма разделяется на частички, обособляющиеся вокруг ядер. Из одной клетки образуется много дочерних. Эта форма размножения обычно чередуется с половой. При почковании на материнской клетке первоначально образуется небольшой бугорок, содержащий дочернее ядро, или нуклеоид. Почка растет, достигает размеров материнской особи и затем отделяется от нее. Эта форма размножения наблюдается у бактерий, дрожжей, сосущих инфузорий, кишечнополостных (гидра). При фрагментации каждая из частей организма достраивает недостающие (плоские черви, кольчецы). При полиэмбрионии эмбрион делится на несколько частей, каждая из которых развивается в самостоятельный организм (у ос наездников, у броненосца). К этой категории относится образование однозиготных близнецов у человека и других млекопитающих. При спорообразовании особь продуцирует специальные клетки бесполого размножения (мхи, папоротники).
Коньюгация — сближение двух соматических клеток и образование между ними цитоплазматического мостика, через который осуществляется обмен наследственным материалом, после чего особи расходятся (у инфузорий), или слияние цитоплазмы обеих клеток (у водорослей, низших грибов). Количество особей при этом не увеличивается, но происходит обновление их наследственного материала. После коньюгации они начинают размножение бесполым путем.
Копуляция — слияние двух одинаковых или разных по форме, размерам и подвижности половых клеток (гамет).
Нерегулярные типы полового размножения: партеногенез, андрогенез, гиногенез.
При партеногенезе новый организм развивается из неоплодотворенной яйцеклетки (пчелы, плоские черви).
Андрогенез (от греч. andros — мужчина) — форма размножения организмов, при которой в развитии зародыша участвует мужское ядро, привнесенное в яйцо сперматозоидом, а женское — не участвует (у наездников, кукурузы, видов табака).
Гиногенез (от греч. gyne — женщина) — форма размножения организма, при которой сперматозоид, проникая в яйцеклетку, стимулирует ее развитие, но ядро его не сливается с ядром яйца и не участвует в последующем развитии зародыша (у некоторых видов нематод, костистых рыб, земноводных и многих покрытосеменных растений).
Образование мужских половых клеток протекает в семенниках (сперматогенез), а женских — в яичниках (оогенез). В гаметогенезе различают четыре периода: размножение, рост, созревание, формирование.
В период размножения первичные половые клетки делятся путем митоза при сохранении диплоидного набора хромосом. В период роста они достигают размеров, характерных для половых клеток каждого вида животных. При созревании первичные половые клетки делятся мейозом, в результате чего образуются гаплоидные половые клетки. Гаплоидные половые клетки в период формирования превращаются в зрелые.
Процессы сперматогенеза и оогенеза в принципе сходны, но между ними есть и различия, в частности, в оогенезе нет периода формирования. В результате сперматогенеза образуются четыре сперматозоида, а оогенез завершается формированием одной яйцеклетки.
Дата добавления: 2014-12-22; просмотров: 2056;