ЦИТОПЛАЗМАТИЧЕСКАЯ НАСЛЕДСТВЕННОСТЬ
Хромосомная теория наследственности установила ведущую роль ядра и находящихся в нем хромосом в явлениях наследственности. Но в то же время уже в первые годы формирования генетики как науки были известны факты, показывающие, что наследование некоторых признаков связано с не хромосомными компонентами клетки (пластидами, митохондриями, хлоропластами) и не подчиняется менделевским закономерностям, основанным на распределении хромосом в ходе мейоза.
Впервые внеядерная наследственность (пестролистность) была описана в 1909 г. Корренсом и Бауэром у растений ночной красавицы (Mirabilis jalapa) и львиного зева, которая обусловлена цитоплазмой. В последующие годы подобные наблюдения были сделаны и на других объектах. Результаты изучения внеядерной наследственности привели к тому, что клетку стали рассматривать как единую целостную систему, определяющую передачу и воспроизведение признаков в потомстве благодаря взаимодействию ядра и цитоплазмы. При удалении ядра клетка погибает, погибает и ядро при выделении его из клетки.
В ядре, точнее в молекулах ДНК, находящихся в хромосомах, вырабатываются те вещества, на основе которых в цитоплазме осуществляется синтез важнейших для жизни клетки соединений – белков, в том числе и ферментов. В цитоплазме же поступающие извне питательные вещества перерабатываются до уровня, необходимого для функций клетки в целом и ее органоидов, в том числе и ядра. Органоиды цитоплазмы – митохондрии – снабжают цитоплазму и ядро необходимой для их жизнедеятельности энергией. Цитоплазма служит барьером, препятствующим проникновению в ядро химических веществ, способных повредить ДНК. Поэтому нарушения, происходящие в цитоплазме, могут оказать влияние на функции как клетки, так и ядра. В свою очередь ядро через вырабатываемые в нем матрицы синтеза белка, оказывает влияние на цитоплазму и происходящие в ней процессы.
Тесную взаимосвязь ядра и цитоплазмы можно наблюдать на примере способности клетки к фотосинтезу. Фотосинтез связан с пластидами и находящимся в них пигментом хлорофиллом. Образование и функции пластид обусловливаются наследственными факторами и действием внешних условий (главным образом света, без которого хлорофилл в пластидах не образуется). Мутации в некоторых локусах хромосом могут частично или полностью нарушать процесс образования пластид и содержащегося в них хлорофилла. Эти хлорофилльные мутации наследуются строго по законам Менделя. Но аномальные (белые) пластиды могут образовываться в клетках, имеющих нормальный набор генов, и при хорошем освещении. Этот признак не наследуется по правилам Менделя. При делении клетки, содержащей такие белые пластиды, образуются дочерние клетки с такими же пластидами, но при скрещивании этот признак передается только по материнской линии и, следовательно, он связан не с хромосомами, а с цитоплазмой. Таким образом, такое важнейшее свойство клетки, как ее способность к фотосинтезу, определяется взаимодействием генов хромосом, структурных элементов цитоплазмы и условий внешней среды.
Исходя из вышеизложенного, система генетического аппарата клетки выглядит так:
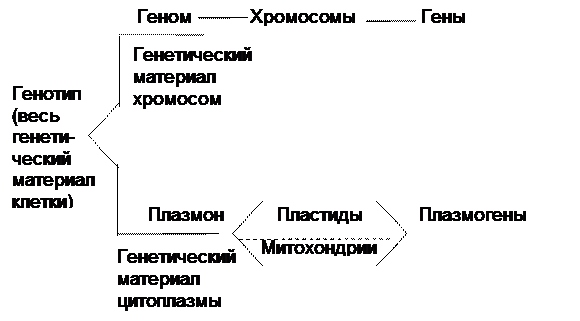 |
Генетическому материалу хромосомного набора (геному) соответствует плазмон, включающий весь генетический материал цитоплазмы. Подобно генам хромосом, в структурных элементах цитоплазмы (пластидах, митохондриях, центросомах, основном ее веществе) находятся материальные носители нехромосомной наследственности. Любая единица цитоплазматической наследственности, соответствующая единице хромосомной наследственности (гену), называется плазмогеном.
Плазмогены могут определять развитие некоторых признаков клетки, обладают способностью удваиваться. Если они утрачиваются клеткой, то хромосомы не могут их воспроизвести, при делении материнской клетки они распределяются между дочерними клетками. Наиболее полно изучены две формы цитоплазматической наследственности: пластидная и цитоплазматическая мужская стерильность (ЦМС).
Пластидная наследственность. Среди органоидов цитоплазмы генетическая непрерывность впервые была установлена для пластид. У многих видов растений встречаются особи, совершенно лишенные окраски (альбиносы) или такие, в листьях которых имеются неокрашенные участки ткани (пестролистность). Альбиносы нежизнеспособны и обычно погибают в фазе проростков. Но отдельные участки ткани без зеленой окраски развиваются в зеленом листе, питаясь за счет нормальных тканей, снабжающих их продуктами фотосинтеза.
У кукурузы, ячменя и некоторых других культур изучены многочисленные хлорофилльные мутации, наследующиеся по законам Менделя. Однако во многих случаях наследование таких изменений не подчиняется менделевским закономерностям, и объяснить его можно, только исходя из представления о генетической непрерывности пластид.
Так, у расы львиного зева (Antirrhinum majus) листья испещрены неправильными желтовато-белыми и зелеными пятнами, причем в одних случаях зеленые пятна на белом фоне, в других - белые пятна на зеленом, и наряду с этим есть целиком зеленые и целиком белые листья. При вегетативном размножении эта раса очень неустойчива; наравне с пестролистными побегами появляются ветви с чисто зелеными листьями и ветви, у которых все листья желтовато-белые, и при дальнейшем вегетативном размножении стойко сохраняют такую окраску.
При половом размножении цветки, возникающие на зеленых ветвях, дают начало семенам, из которых образуются зеленые растения. Цветки на белых ветвях завязывают семена, дающие начало сеянцам с белыми листьями, которые быстро гибнут из-за недостатка пищи. И наконец, на пестролистных ветвях цветки завязывают семена, из которых образуются как сеянцы с зелеными и белыми листьями, так и пестролистные. Для формирования окраски листьев семенного потомства решающее значение имеет окраска листьев побегов, на которых образовались семена.
При опылении цветков зеленых побегов пыльцой с цветков белых побегов все потомство имеет зеленые листья, а при обратном скрещивании – белые. Таким образом, окраска листьев в этом случае наследуется по женской линии, и характер потомства определяется исключительно материнским растением. Пыльца может быть любой, и ее происхождение никак не отражается на признаках потомков:
Мать зеленолистная
| ♀ зел. × ♂ зел. | ♀ зел. × ♂ пестрол. | ♀ зел. × ♂ бел. |
| F1 зел. | F1 зел. | F1 зел. |
Мать пестролистная
| ♀ пестр. × ♂ пестр. | ♀ пестр. × ♂ зел. | ♀ пестр. × ♂ бел. |
| F1 пестролистные, зеленые, белые | F1 пестролистные, зеленые, белые | F1 пестролистные, зеленые, белые |
Мать белолистная
| ♀ бел. × ♂ зел. | ♀ бел. × ♂ пестр. | ♀ бел. × ♂ бел. |
| F1 белые | F1 белые | F1 белые |
Такая картина наследования стойко повторяется из поколения в поколение. Объясняется этот тип наследования тем, что пластиды обладают генетической непрерывностью, причем зеленые дают начало зеленым, а белые – белым. Воспроизводство пластид происходит независимо от деления клетки и при митозе они распределяются между дочерними клетками число случайно. Поэтому, если клетка содержит смесь зеленых и белых пластид, то в результате митоза получаются как клетки со смешанными пластидами, так и либо с одними зелеными, либо с одними белыми пластидами:
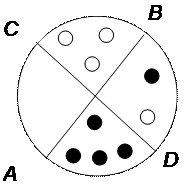
Если клеточная перегородка пройдет по линии А – В, то одна дочерняя клетка будет содержать только белые пластиды, другая – зеленые и белые. Если же клеточная перегородка пройдет по линии С – D, то одна дочерняя клетка будет содержать только зеленые пластиды, а другая – смесь белых и зеленых. В дальнейшем из этих клеток образуются зеленые, белые и мозаичные растения. Во время макро- и микроспорогенеза пластиды попадают в яйцеклетки, но не в пыльцевые зерна, что обусловливает чисто материнское их наследование.
Нормальное материнское растение дает только нормальное потомство, а аномальное – только аномальное, независимо от фенотипа отцовской формы. Наследственность, передаваемая только через мать, называется материнской. Материнская наследственность характерна только для тех признаков, на которые влияют цитоплазма и ее органоиды. Наследование признаков через цитоплазму получило название цитоплазматической наследственности.
В процессе изучения роли цитоплазмы в наследственности возникают осложнения, затрудняющие правильное решение этого вопроса и являющиеся следствием трех причин.
1. Иногда в цитоплазме находятся инородные, органически с ней не связанные включения, чаще вирусы, реже – бактерии; их присутствие в клетке ведет к изменению жизнедеятельности и развитию определенных признаков. Вместе с цитоплазмой яйцеклетки включения попадают в зиготу, вызывая аналогичные материнскому организму отклонения потомства от нормы и создавая картину наследования их через цитоплазму. Примером такой наследственности служит передача у дрозофилы по типу материнской наследственности высокой чувствительности к двуокиси углерода (СО2). Скрещивание чувствительных к СО2 самок с нормальными самцами дает чувствительное потомство, и этот признак стойко передается, если гибридные самки скрещиваются с нормальными самцами. Самки, происходящие от исходной чувствительной прародительницы, продолжают производить чувствительное потомство, даже в том случае, когда с помощью возвратных скрещиваний все их хромосомы заменены хромосомами нормальной линии. Скрещивание нормальных самок с чувствительными самцами показало, что чувствительность может передаваться и с мужской стороны, но с малой частотой, причем соотношение чувствительных и нормальных потомков не укладывается в закономерности хромосомной наследственности. Все эти данные подтверждают, что чувствительность к СО2 наследуется у дрозофилы через цитоплазму.
В течение длительного времени оставалось неизвестным, что же представляет собой цитоплазматический фактор, обусловливающий чувствительность дрозофилы к СО2 и передающийся из поколения в поколение. Лишь в последние годы удалось с помощью новейших лабораторных методов выделить из дрозофилы и изучить фактор чувствительности к СО2, оказавшийся типичным вирусом. Этот вирус размножается только в цитоплазме клеток дрозофилы и делает нормальных мух чувствительными к СО2.
2. Некоторые гены, находящиеся в хромосомах, вызывают изменение цитоплазмы, в том числе и яйцеклетки, в результате чего такая цитоплазма оказывает влияние на первые стадии развития зиготы, направляя его по типу материнского организма.
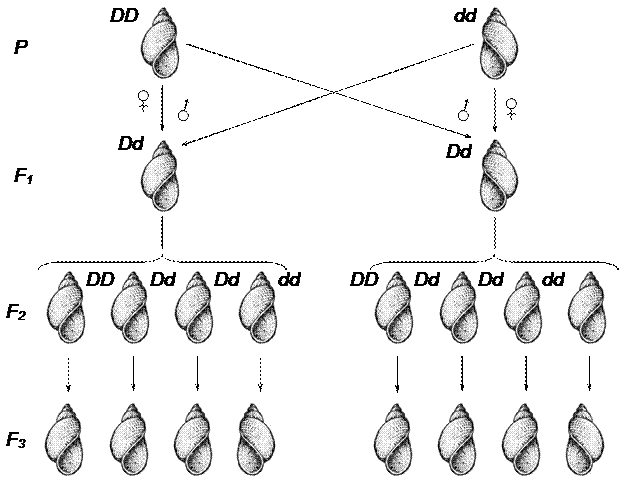
Ярким примером материнской наследственности, вызванной изменением цитоплазмы яйцеклетки ядром матери, служит наследование извитости раковины у некоторых линий пресноводной улитки (Limnaea). У некоторых линий раковина образует завиток по часовой стрелке (декстральный), у других линий этот завиток направлен против движения часовой стрелки (синистральный). Декстральный (правозакрученный) завиток контролируется доминантной аллелью (D), синистральный (левозакрученный) – рецессивным аллелем (d). При реципрокных скрещиваниях улиток этих линий раковина у F1 всегда бывает того типа, какой был у матери.
РР ♀ DD × ♂ dd РР ♀ dd × ♂ DD
завиток завиток завиток завиток
декстральный синистральный синистральный декстральный
F1 Dd F1 Dd
завиток декстральный завиток синистральный
Благодаря тому, что улитки являются гермафродитами, они могут размножаться как путем самооплодотворения, так и путем скрещивания.
От самооплодотворения гетерозиготных форм F1 (Dd) в обоих скрещиваниях все потомки F2 обладают декстральным завитком (правозакрученным), хотя гибриды F1 (материнские формы) различались по фенотипу. Когда было исследовано потомство от каждой особи в отдельности, то выяснилось, что ¼ семей имели левый завиток, ¾ - правый. Простое менделевское расщепление по данной паре признаков выявилось не в F2, а только в F3. При этом типе наследования фенотип потомков соответствует генотипу матери, а не генотипу зигот, из которых они развиваются. Это может быть только при допущении, что данный признак предопределяется генотипом материнского организма в цитоплазме яйца в процессе его развития (рис. 41).
Рис. 41. Наследование извитости раковины у моллюска
 |
Рассмотренный тип наследования и является в собственном смысле слова материнским. Как было выяснено, направление завитка раковины определяется характером спирального дробления оплодотворенного яйца, т. е. расположением бластомеров по спирали вправо или влево, что в свою очередь зависит от ориентации веретена при втором делении дробления. Проявление признака – направление закручивания раковины у прудовика – как бы отстает на поколение от генотипической формулы. Решающую роль в таком отставании играет генотип матери, определяющий прежде всего свойства цитоплазмы яйцеклетки, от которого и зависит направление закручивания раковины. Такое явление получило название предетерминации цитоплазмы генотипом матери, или материнского эффекта.
3. Непосредственное влияние материнского организма на развитие зародыша также нередко ведет к известному сходству потомства с матерью, хотя оно и не обусловлено наследственностью. Мать оказывает на потомство большее влияние, чем отец, благодаря тому, что условия эмбрионального развития потомства почти целиком зависят от материнского организма, обеспечивающего эмбрион питательными веществами, будь то яйцо птицы, икра рыбы или эмбрион млекопитающих. Влияние материнского организма особенно явно выступает в реципрокных скрещиваниях при получении межвидовых гибридов. Так, при скрещивании двух видов кипрея – Epilobium hirsutum и E. luteum – хорошо заметна разница в плодовитости гибридов.
Если материнской формой служит E. hirsutum, а отцом - E. luteum, то F1 характеризуется резким угнетением роста и совершенно стерильной пыльцой. Обратное скрещивание ♀ E. luteum × ♂ E. hirsutum дает нормально развитое плодовитое потомство. Такие гибриды в течение ряда поколений опыляли пыльцой E. hirsutum. С каждым поколением потомки становились все более похожими на отцовский вид и после 14 таких возвратных скрещиваний стали фенотипически неотличимыми от E. hirsutum вследствие полной замены хромосом от E. luteum на хромосомы E. hirsutum. Однако их цитоплазма осталась от вида E. luteum, так как она передается только яйцеклетками, то есть только с материнской стороны. Опыление таких гибридов 14 поколения пыльцой E. hirsutum давало точно такой же результат, какой получили в F1 от скрещивания ♀ E. luteum × ♂ E. hirsutum – потомки были крайне низкорослыми и имели стерильную пыльцу. Это значит, что задержка роста таких межвидовых гибридов и стерильность их пыльцы обусловлены взаимодействием хромосомных генов E. hirsutum с какими-то наследственными факторами, локализованными в цитоплазме E. luteum. Природа этих плазмогенов пока неизвестна.
Дата добавления: 2019-07-26; просмотров: 1571;