НАСЛЕДОВАНИЕ ПРИЗНАКОВ ПРИ СЦЕПЛЕНИИ ГЕНОВ
Используя принцип генетического анализа, мы в ранее изложенных разделах моно- и дигибридного скрещивания убедились, что законы Менделя действуют в тех случаях, когда признаки и свойства организмов определяются генами, расположенными в разных парах хромосом. Число признаков и свойств организмов, контролируемых генами, очень велико, а число пар хромосом, характерное для каждого биологического вида, относительно мало и постоянно.
Если предположить, что в каждой хромосоме не один ген, а два, три и более, то тогда следует признать, что третий закон Менделя (о независимом наследовании генов) касается распределения хромосом, а не генов. Гены, расположенные в разных парах хромосом, называются несцепленными, и развернутые формулы, например, дигибрида выглядят таким образом: 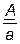
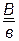 или
или 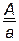
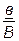 . Черточки символизируют хромосомы. Гены, вместе с их носителями – хромосомами закономерно распределяются в мейозе при образовании гамет, свободно и независимо комбинируются и обусловливают расщепление в F2 по фенотипу в отношении 9 : 3 : 3 : 1.
. Черточки символизируют хромосомы. Гены, вместе с их носителями – хромосомами закономерно распределяются в мейозе при образовании гамет, свободно и независимо комбинируются и обусловливают расщепление в F2 по фенотипу в отношении 9 : 3 : 3 : 1.
Теперь представим себе, что две аллельные пары Аа и Вв находятся не в двух разных хромосомах, а в одной гомологичной паре хромосом: при написании генотип дигибрида обозначим следующим образом:  или
или 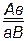 . Гены различных локусов, расположенных в одной хромосоме, называются сцепленными. В одной хромосоме могут располагаться не только два, но и три, четыре и более неаллельных генов. Формулу гетерозиготного состояния по трем, например, генам можно написать следующим образом:
. Гены различных локусов, расположенных в одной хромосоме, называются сцепленными. В одной хромосоме могут располагаться не только два, но и три, четыре и более неаллельных генов. Формулу гетерозиготного состояния по трем, например, генам можно написать следующим образом: 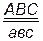 ,
, 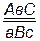 ,
, 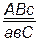 . Гены, расположенные в одной паре гомологичных хромосом, наследуются целой группой, образуя одну группу сцепления. Групп сцепления в организме столько, сколько хромосом в гаплоидном наборе. Например, у кукурузы 2n=20, групп сцепления у нее – 10 (n=10), у гороха 2n=14, групп сцепления – 7 и т. д.
. Гены, расположенные в одной паре гомологичных хромосом, наследуются целой группой, образуя одну группу сцепления. Групп сцепления в организме столько, сколько хромосом в гаплоидном наборе. Например, у кукурузы 2n=20, групп сцепления у нее – 10 (n=10), у гороха 2n=14, групп сцепления – 7 и т. д.
Причем гены одной группы сцепления наследуются независимо от другой. Это вносит ограничения в третий закон Менделя (о независимом комбинировании признаков у гибридов). Здесь гены наследуются совместно.
Существует различная степень сцепления между генами одной и той же группы сцепления. Крайними являются случаи, когда гены всегда или почти всегда наследуются совместно (полное сцепление). Наоборот, есть случаи, когда возможность их совместного наследования чуть больше, нежели порознь.
Проанализируем характер расщепления гибридов при полном сцеплении генов. Для этого необходимо скрещивание двух гомозиготных организмов:
РР ♀ 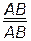 × ♂
× ♂ 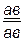
гаметы: ♀ АВ ♂ ав
F1 .
Для получения F2 необходимо скрестить между собой гибриды первого поколения:
РР ♀F1 × ♂ F1 .
Родители будут давать только по два типа гамет (при условии полного сцепления) в равном соотношении.
гаметы: ♀ АВ, ав ♂ АВ, ав
F2 1 ; 2  ; 1 .
; 1 .
Проведя анализирующее скрещивание дигибрида, получим:
РР ♀ F1 × ♂
гаметы: ♀ АВ, ав ♂ ав
Fан 1 ; 1 .
Расщепление по генотипу в F2 соответствует 1 : 2 : 1, а по фенотипу (при полном доминировании) – 3 : 1. В потомстве анализирующего скрещивания наблюдается расщепление 1 : 1. Из этого следует, что не произошло никаких перегруппировок признаков, которые могли бы привести к появлению новых их комбинаций. Другими словами, сцепленные гены наследуются в потомстве как пара аллельных генов.
Однако полное сцепление генов в природе встречается редко. Оно отмечено у самок тутового шелкопряда, у самцов мушки-дрозофилы, когда гетерозиготы дают всего лишь два типа гамет. У большинства видов растений и животных сцепление генов неполное, в этом случае сцепленные гены часто наследуются вместе.
Например, при скрещивании двух сортов гороха, один из которых имел морщинистые семена и растения без усов, а другой – гладкие семена и растения с усами, получили гибриды, растения которых имели усы и гладкие семена. В результате анализирующего скрещивания дигибрида образовалась 4 фенотипических класса с таким соотношением:
516 растений с гладкими семенами и усами,
492 растения с морщинистыми семенами и без усов,
9 растений с гладкими семенами и без усов,
7 растений с морщинистыми семенами и с усами.
Эти результаты свидетельствуют о том, что мы имеем дело с неполным сцеплением генов, так как дигибрид образует не два, а четыре типа гамет. Два типа гамет образуется, если сцепление генов полное, а два – если это сцепление нарушается. Нарушается сцепление между генами в результате обмена между гомологичными участками гомологичных хромосом. Это явление называют кроссинговером. Кроссинговер является результатом перекреста хромосом, поэтому его называют еще перекрестом (Crossing over – образование перекреста).
Вернемся к нашему примеру и рассмотрим его на уровне генных формул.
|
|
× ♂
гаметы: ♀ ав ♂ АВ
|
.
|
|
× ♂
гаметы: ♀ АВ, ав, Ав, аВ, ♂ ав
Fан ; ; 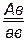 ;
; 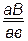 .
.
|
|
|
|
Как видим, в результате анализирующего скрещивания не обнаружили расщепление 1 : 1 : 1 : 1, как было при независимом наследовании признаков. Два преобладающих класса (516 и 492), где сцепление генов не претерпело кроссинговера, похожи на родителей и их называют некроссоверными. Два других класса (9 и 7) с новой комбинацией генов, претерпевших кроссинговер, называют кроссоверными. Это свидетельствует о том, что дигибрид давал разные гаметы – кроссоверные и некроссоверные. Гаметы АВ и ав, имеющие такое же сочетание генов, как у родителей, называются некроссоверными. Гаметы Ав и аВ, имеющие иное сочетание генов, чем у родителей, являются кроссоверными. Поскольку сцепленные гены имеют тенденцию к совместному наследованию, то кроссоверных гамет и особей всегда будет меньше, чем некроссоверных. Так в нашем случае некроссоверных особей было 1008 (516+492), а кроссоверных – 16 (9+7).
Таким образом, кроссинговер происходил с частотой: 1 кроссоверная особь на 63 некроссоверных (1008 : 16 = 63). Ввиду того, что при кроссинговере между родительскими хромосомами осуществляется взаимный обмен, это и вынуждает подсчитывать кроссоверные классы вместе как результат одного события.
Мы рассмотрели пример, когда рецессивные гены переданы гибриду от одного родителя. Но могут быть случаи, когда рецессивное гены передаются гибриду от двух родителей.
|
|
 × ♂
× ♂ 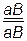
гаметы: ♀ Ав ♂ аВ
|
В результате анализирующего скрещивания было получено:
|
|
× ♂
гаметы: ♀ Ав, аВ, АВ, ав, ♂ ав
Fан ; ; ;
285 283 4 5
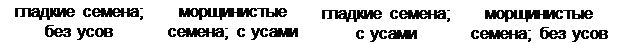 |
Некроссоверных особей оказалось 568 (285+283), кроссоверных 9 (4+5). И в этом случае одна кроссоверная особь приходится на 63 некроссоверных (568:9=63,1). Здесь прослеживается закономерность, указывающая на то, что кроссинговер происходит с определенной частотой, не зависящей от того, в каких комбинациях особь, образующая гаметы, получила рассматриваемые неаллельные гены.
Генетически обнаружить кроссинговер можно по перекомбинации признаков в потомстве, которое можно определить, получив F2 или путем анализирующего скрещивания. Поскольку анализ F2 более сложный, чаще используют анализирующее скрещивание дигибрида. Другими словами, кроссинговер можно обнаружить, если гены находятся в гетерозиготном состоянии, то есть или . При гомозиготном состоянии генов ( , , 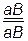 , ) выявить это нельзя, так как обмен идентичными участками не дает новых комбинаций генов в гаметах и потомстве. Наличие такого механизма обмена между родительскими парами, то есть рекомбинации генов, расширяет возможности комбинативной изменчивости в эволюции.
, ) выявить это нельзя, так как обмен идентичными участками не дает новых комбинаций генов в гаметах и потомстве. Наличие такого механизма обмена между родительскими парами, то есть рекомбинации генов, расширяет возможности комбинативной изменчивости в эволюции.
Величину кроссинговера можно учесть. Она измеряется отношением числа кроссоверных особей в потомстве анализирующего скрещивания к общему числу особей, принятому за 100%.
В нашем примере (1 случай) общее число особей 1024 (1008+16) берут за 100 %, а число кроссоверных особей (16) – за х %.
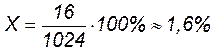 .
.
Величина перекреста хромосом отражает силу сцепления генов в хромосоме: чем больше величина перекреста, тем меньше сила сцепления. Т. Морган предположил, что частота кроссинговера показывает относительное расстояние между генами: чем чаще осуществляется кроссинговер, тем дальше отстоят друг от друга гены в хромосоме, чем реже кроссинговер, тем ближе они друг к другу. На основе многочисленных генетических исследований Т. Морган выдвинул гипотезу линейного расположения генов в хромосоме. Один процент перекреста принят за условную единицу силы сцепления или относительного расстояния между генами. В русской литературе эту единицу называют морганидой (в честь Т. Моргана). Из нашего примера следует, что гены А и В сцеплены и находятся на расстоянии 1,6 кроссоверных единиц (морганид).
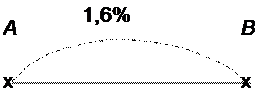
Перекрест между хромосомами может быть одинарным, если он происходит в одном месте, двойным – в двух местах одновременно, тройным – в трех точках и т. д., т. е. кроссинговер может быть множественным. Двойной кроссинговер – это обмен средним участком гомологичных хромосом. Между близко расположенными генами двойных кроссинговеров не происходит, так как уже осуществившийся перекрест препятствует возникновению новых в близлежащих участках хромосомы.
В том случае, когда процент кроссинговера достигает 50, мы не можем определить, имеем ли мы дело со сцеплением анализируемых генов или с независимым их расщеплением. При таком проценте кроссинговера четыре типа гамет, образуемых дигибридом, дают при возвратном скрещивании соотношение 1 : 1 : 1 : 1, т. е. то отношение, какое ожидается при независимом расщеплении. Иными словами, непосредственное измерение величины кроссинговера между любыми двумя локусами не может превышать 50%, но общая протяженность расстояния между ними на карте хромосомы может превышать эту величину.
Обычно при кроссинговере происходит комплементарный обмен строго идентичными участками хроматид гомологичных хромосом. Однако у ряда организмов установлен факт обмена неравными участками (неравный кроссинговер).
Кроссинговер характерен для мейотического деления, однако как исключение может встречаться и в митотических делениях соматической ткани. Такой кроссинговер называется митотическим, или соматическим. Соматический кроссинговер возможен и в половых хромосомах, и в аутосомах.
Дата добавления: 2019-07-26; просмотров: 1571;