Взаимодействие неаллельных генов
1. Комплементарность – тип взаимодействия неаллельных генов, при котором доминантные гены из разных аллельных пар сочетаясь вместе в генотипе обеспечивают появление нового признака (поэтому генотип А-В- всегда имеет новый фенотип).
2. Эпистаз - тип взаимодействия неаллельных генов, при котором гены одной аллельной пары подавляют действие генов другой аллельной пары. Различают доминантный и рецессивный эпистаз.
3. Полимерия - тип взаимодействия неаллельных генов, при котором гены из разных аллельных пар отвечают за одно выражение признака. При этом они обозначаются одинаковыми буквами, но с разными индексами: А1А1А2А2; а1а1а2а2.
При кумулятивной полимерии степень выраженности признака зависит от числа доминантных генов в генотипе: чем больше доминантных генов, тем признак более выражен.
При некумулятивной полимерии наличие хотя бы одного доминантного гена из любой пары обеспечивает сразу максимальное выражение признака.
Аутосомное наследование
Аутосомное наследование – это наследование признаков, за развитие которых отвечают гены, расположенные не в половых хромосомах (аутосомах). Аутосомное наследование может проходить по доминантному и рецессивному типу.
Наследование, сцепленное с полом
Наследование, сцепленное с полом — наследование признаков, за развитие которых отвечают гены, расположенные в половых хромосомах.
Пол, несущий одинаковые половые хромосомы (ХХ), называется гомогаметным и образует один тип гамет.
Пол, несущий разные половые хромосомы (ХУ), называется гетерогаметным и образует 2 типа гамет.
При наследовании, сцепленным с Х-хромосомой, гомогаметный пол может быть представлен гомозиготой (доминантной - ХАХА или рецессивной - ХаХа) или гетерозиготой — ХАХа. Гетерогаметный пол всегда представлен гемизиготой — ХАУ или ХаУ
У некоторых организмов (например, млекопитающие) гомогаметный пол — женский, у других организмов (например, бабочки, птицы) гомогаметный пол мужской.
При наследовании, сцепленным с У-хромосомой, признак передается только по линии гетерогаметного пола. Такое наследование называется голандрическим.
Генетика пола
·У некотрых организмов пол определяется составом половых хромосом: у млекопитающих гомогаметный пол - женский, у других организмов (например, бабочки, птицы) гомогаметный пол мужской.
·У некоторых организмов пол определяется числом половых хромосом. Например, у кузнечиков самки - ХХ, а самцы - Х0.
·У дрозофилы наследование пола зависит не столько от числа Х-хромосом, сколько от соотношения числа Х-хромосом к числу наборов аутосом. Такое отношение называют половым индексом. Для генотипа ХХ: 2Х:2А=1, половой индекс равен 1, то развивается самка. Для генотипа ХУ: 1Х:2А=0,5 — развивается самец. При промежуточном соотношении развиваются интерсексы (2Х:3А=0,67), то есть мухи, имеющие промежуточный фенотип.
·У некоторых организмов (пчел, муравьев, ос) существует гапло-диплоидный тип определения пола. У этих особей нет половых хромосом. Самки развиваются из оплодотворенных яиц, а самцы из неоплодотворенных.
·У некоторых организмов в определении пола большую роль играет внешняя среда.
Хромосомная теория наследственности
В 1908-1911 гг. американским генетиком Т.Морганом был установлен ряд закономерностей наследования, получивший впоследствии названиехромосомной теории наследственности.
Основными положениями этой теории являются следующие:
1) гены расположены в хромосомах, представляющих собой группу сцепления генов; число групп сцепления генов равно гаплоидному набору хромосом, специфическому для каждого вида организмов;
2) гены расположены в пределах хромосомы линейно; место, занимаемое геном в хромосоме, называется «локус».
3) сцепление генов может нарушаться в результате кроссинговера (перекреста хромосом), в ходе которого гомологичные хромосомы обмениваются между собой одним или несколькими генами;
4) частота обмена генами (частота кроссинговера) пропорциональна расстоянию между генами в пределах хромосомы.
Изучение частоты (процента) кроссинговера между генами, локализованными в пределах одной хромосомы, позволяет установить относительное расстояние между ними, оцениваемое в единицах морганидах (1 морганида равна расстоянию между генами с частотой кроссинговера в 1%). На основе многочисленных экспериментов по изучению кроссинговера составляются генетические карты хромосом. В настоящее время они составлены для всех хромосом мухи-дрозофилы, отдельных хромосом томатов, кукурузы и других видов.
Биологическая роль хромосом
Хромосомы во взаимодействии с внехромосомными элементами клетки обеспечивают:
1) хранение генетической наследственной информации;
2) использование генетической информации для создания и поддержания клеточной организации;
3) регуляцию «считывания» генетической информации;
4) удвоение генетического материала;
5) перекомбинирование генетического материала в процессе мейоза;
6) передачу генетического материала от материнской клетки дочерним.
Сцепленное наследование
Сцепленное наследование – наследование признаков, за которые отвечают гены, расположенные в одной хромосоме.
Изучал Т. Морган. При скрещивании дигетерозиготных самок дрозофил (АаВв), имеющих серое тело и нормальные крылья (доминантные признаки) с самцами дрозофил (аавв), имеющими черное тело и укороченные крылья (рецессивные признаки) в потомстве по законам Менделя ожидалось расщепление по фенотипу и генотипу как в анализирующем скрещивании 1:1:1:1. Однако Морганом было получено расщепление:
- 41,5% - с серым телом, нормальными крыльями;
- 41,5% - с черным телом, укороченными крыльями;
- 8, 5% - с серым телом, укороченными крыльями;
- 8,5% - с черным телом, нормальными крыльями.
Морган делает вывод, что гены, отвечающие за окраску тела и длину крыльев, расположены в одной хромосоме и поэтому наследуются сцеплено. Наличие фенотипических групп, отличных от родительского организма, является результатом кроссинговера.
Сцепление генов может быть полным или частичным.
При полном сцеплении кроссинговер между генами по каким-либо причинам невозможен, или гены настолько близко расположены друг к другу, что вероятность кроссинговера между ними настолько мала, что им можно пренебречь.
Например, и дигетерозиготных самцов дрозофил, имеющих серое тело и крылья нормальной длины кроссинговер не возможен из-за отсутствия фермента, обеспечивающего конъюгацию хромосом.
При частичном сцеплении между генами возможен кроссинговер, и необходимо учитывать процент образования кроссоверных гамет.
Особенности взаимосвязи
между генотипом и фенотипом
Генотипом называется совокупность всех генов данного организма. Фенотип - это совокупность всех внешних и внутренних признаков организма. Между генотипом и фенотипом существуют следующие взаимосвязи (взаимоотношения).
1. Ген контролирует (кодирует) структуру белка одного вида, обладающего, чаще всего, ферментативной активностью. Регулируя биосинтез белков или другие важные биохимические превращения, фермент оказывает влияние на проявление того или иного признака и, тем самым, на формирование или изменение фенотипа организма.
2. Однако проявление действия генов и характер возникающего признака зависят от условий окружающей среды. Условия среды воздействуют на особенности протекания и скорость биохимических реакций и, тем самым, оказывают влияние на фенотипический признак или степень его выраженности. Таким образом, в определенных пределах (пределах нормы реакции) может происходить изменение фенотипа без предварительного изменения генотипа. Это явление получило название модификационной (фенотипической) изменчивости. Пределы таких независимых изменений фенотипа, тем не менее, контролируются генотипом.
3. Изменения фенотипа, обеспечивающие адаптацию организма к конкретным условиям среды, происходящие независимо от генотипа, не наследуются потомством, и, казалось бы, не имеют эволюционного значения. Однако, любые адаптации организма (изменения фенотипа), повышающие уровень его приспособленности к окружающей среде, повышают, тем самым, и его шансы на репродуктивный успех. В таком случае генотипы этих организмов скорее сохранятся в эволюционной линии (цепочке последующих поколений), чем будут устранены из нее. Следовательно, отдельные ненаследуемые (фенотипические) изменения обеспечивают выживание организмов (генотипов) – обладателей аллелей (генов), контролирующих другие адаптивные признаки, информация о которых наследуется потомством и примет дальнейшее участие в эволюции.
Ген как единица наследственности
Согласно современным представлениям, ген - это участок молекулы ДНК, кодирующий иРНК (а, следовательно, первичную структуру белка), или тРНК, или рРНК.
Вся совокупность генов составляет генотип организма.
Свойства генов:
1. Способность к самоудвоению.
2. Способность мутировать независимо от остальных генов, дискретно изменяя своё внутреннее состояние как целое, т.е. порождая аллели.
3. Способность к гомологичному синапсису при коньюгации в мейозе.
4. Неделимость и несмешиваемость в процессах коньюгации и кроссинговера.
5. Устойчивость локализации в геноме относительно других генов.
6. Способность контролировать развитие некоторого признака или небольшого их числа, что отражено в названиях генов.
7. Высокая устойчивость к мутациям и другим изменениям.
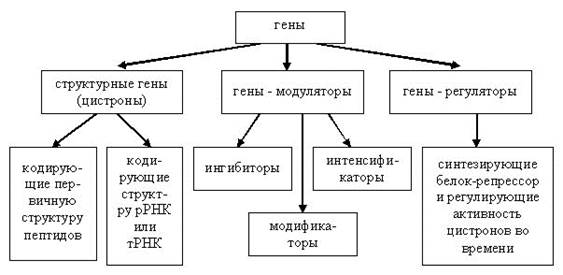
Классификация генов:
1. Структурные гены контролируют развитие конкретных признаков. Каждый ген контролирует синтез фермента, регулирующего определённый этап метаболизма.
2. Гены-модуляторы смещают в ту или другую сторону процесс развития признака, кодируемого структурным геном (цистроном).
а) гены-ингибиторы могут тормозить развитие отдельных признаков, обусловливаемых плейотропным действием структурных генов, или даже целиком подавлять функции других генов
б) гены-интенсификаторы усиливают функцию цистронов
в) гены-модификаторы оказывают влияние на степень проявления признака, обусловливаемого расположенным в другом локусе структурным геном.
3. Гены-регуляторы координируют активность генов, регулируя «включение-выключение функции» различных генов во времени в процессе онтогенеза.
Генотип как целостная исторически сложившаяся система взаимодействующих клеток
Генотип формируют все (ядерные и внеядерные) гены данного организма. Несмотря на то, что гены локализованы в различных самостоятельных структурах (хромосомы, кольцевые молекулы ДНК митохондрий и пластид) все они взаимосвязаны и могут взаимодействовать друг с другом в процессе развития организма (формирования фенотипа). Поэтому вся совокупность генов организма представляет собой взаимодействующую систему организма – генотип.
Как любая система, генотип характеризуется целостностью и единством: повреждение (устранение) даже одного гена нарушает свойства всей системы, что может проявиться в резком снижении жизнеспособности или даже гибели организма. Генотип формируется в ходе эволюции, в процессе которой структура генов изменяется. Поэтому генотип рассматриваетсякак результат процесса эволюции, или исторически сложившаяся система взаимодействующих генов.
Различают аллельные и неаллельные взаимодействия генов.
Составляющие генотип гены могут обладать различными свойствами:
· структурные гены кодируют структуру молекулы и-РНК (а в конечном итоге структуру белковой молекулы одного вида), транспортной или рибосомной РНК (т-РНК, р-РНК);
· гены-регуляторы осуществляют активизацию или угнетение активности структурных генов, за счет синтеза белка-репрессора;
· ген-промотор обеспечивает инициацию процесса транскрипции (синтеза и-РНК), это место прикрепления РНК-полимеразы.;
· ген-оператор включает в работу или выключает из нее структурный ген. Если ген-оператор заблокирован белком репрессором, то транскрипция со структурных генов не происходит. Если ген оператор свободен от белка репрессора, РНК-полимераза может вести транскрипцию со структурных генов.
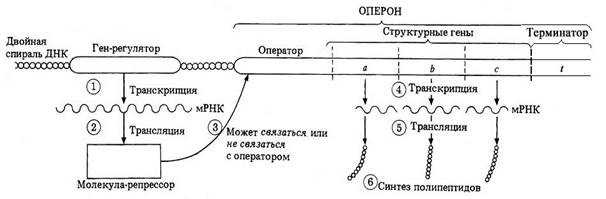
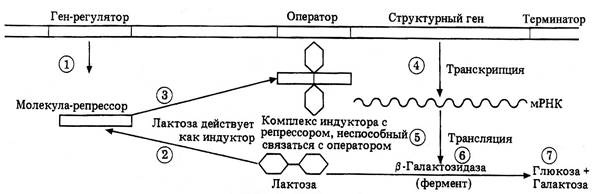
Регуляция активности генов на уровне транскрипции (на примере лактозного оперона)
Единицей транскрипции является транскриптон, или оперон (термин «оперон» введён Ф. Жакобом и Ж. Моно в 1961 году). Оперон - это участок ДНК, транскрипция которого приводит к образованию молекулы иРНК. Структура оперона изучена подробно пока только у прокариот. Оперон может состоять из одного, двух и более тесно сцепленных структурных генов, кодирующих белки (ферменты), а также регуляторных элементов. Участком начала транскрипции является промотор, состоящий из нескольких десятков нуклеотидов ДНК, с которым специфически связывается осуществляющий транскрипцию фермент РНК-полимераза. От промотора зависит, к какой из двух цепей ДНК присоединится РНК-полимераза и начнётся транскрипция. Транскрипция происходит только на одной («лидирующей») цепочке ДНК, и химическая структура промотора определяет эту цепь.
В состав оперона входит также ген-оператор длиной в несколько десятков нуклеотидов, с которым связывается репрессор (специфический белок), обусловливающий отрицательную регуляцию: в этом случае ДНК-полимераза не может перемещаться вдоль оперона и транскрипция структурных генов не происходит. Если же оператор не связан с репрессором, то РНК-полимераза, смещаясь вдоль структурных генов (цистронов), транскрибирует их.
Репрессор, контролирующий транскрипцию оперона, кодируется геном-регулятором, который не обязательно входит в состав оперона: он может находиться за пределами оперона этой же хромосомы или в иной хромосоме. Один репрессор может контролировать транскрипцию нескольких оперонов. Молекула репрессора имеет участок узнавания эффектора, который активирует, или, наоборот, инактивирует репрессор, связываясь с ним. Оперон заканчивается терминатором, представляющим собой один из трёх триплетов, не кодирующих аминокислоты (УАА, УАГ, УГА). Синтезируемая сразу на нескольких цистронах общая цепочка РНК распадается на фрагменты - иРНК, соответствующие каждому отдельно взятому цистрону.
Включение оперона происходит при проникновении в цитоплазму субстрата (например, лактозы), для химического превращения которого требуется соответствующий фермент. Субстрат, соединяясь с репрессором, лишает последний возможности блокировать ген-оператор. Тогда происходит транскрипция структурного гена и синтезируется необходимый белок. Фермент вызывает вовлечение субстрата в процессы клеточного метаболизма. В ходе последнего количество субстрата уменьшается, что приводит к высвобождению репрессора, блокированию оператора и прекращению транскрипции.
|
|
Виды изменчивости
Изменчивость – способность организма приобретать новые признаки в процессе индивидуального развития.
Различают: 1) наследственную изменчивость (генотипическую): мутационную и комбинативную; 2) ненаследственную изменчивость (фенотипическую) – модификационную.
Модификационная изменчивость – изменчивость, затрагивающая только фенотип и не затрагивающая генотип.
Особенности:
1. Не наследуется (так как не затрагивает генотип).
2. Носит групповой характер - сходные изменения возникают у группы особей.
3. Предсказуема – результат действия фактора можно предсказать.
4. Направлена – изменения, возникающие под действием фактора, часто носят адаптивный характер.
5. Обратима – возникающие изменения могут быть обратимы. Однако если действие фактора не специфично или он действует в критический период развития, то могут возникать необратимые изменения – морфозы.
6. Границы изменчивости признака называются нормой реакции и определяются генотипом.
Модификационную изменчивость изучают с помощью вариационно-статистического метода. При этом конкретные значения признака – варианты – размещают в порядке возрастания значений, образуя вариационный ряд. Учитывая встречаемость отдельных вариант, или средних значений классов, составляют вариационные кривые. При этом по оси Х откладывают значения вариант, а по оси У – частоту их встречаемости. Установлено, что вариационная кривая имеет вид перевернутой параболы.
Значение: обеспечивает приспособленность организма к изменяющимся условиям существования, что способствует выживаемости организмов, а значит их успеху в эволюции в целом.
Комбинативная изменчивость – не связана с изменениями генов, а только с их перекомбинацией у потомков. Реализуется в ходе полового размножения.
Причины:
1) кроссинговер;
2) независимое расхождение гомологичных хромосом в анафазу I мейоза, имеющее место при образовании гамет;
3) явление случайного оплодотворения.
Значение:
1) способствует увеличению генетического разнообразия потомства;
2) лежит в основе гибридологического метода в генетике и селекции, позволяет получать организмы с необходимыми человеку комбинациями признаков.
Мутационная изменчивость – вызывается мутациями – непредсказуемыми скачкообразными изменениями генотипа (а именно: генома, хромосом или генов).
Особенности:
1. Наследуется, так как затрагивает генотип.
2. Носит индивидуальный характер.
3. Непредсказуема – невозможно предвидеть какие изменения возникнут на действие фактора.
4. Ненаправлена – изменения не носят приспособительный характер.
Классификации мутаций:
1. По изменению генотипа: генные, хромосомные, геномные.
2. По влиянию на жизнеспособность: летальные, полулетальные, нейтральные.
3. По поведению в гетерозиготе: доминантные и рецессивные.
4. По отношению к генеративному пути: соматические (возникают в обычных клетках тела и ненаследуются) и генеративные (в половых клетках, поэтому наследуются).
5. По локализациив клетке: ядерные (в ДНК ядра) и цитоплазматические (в ДНК митохондрий и пластид).
6. По причине, вызывающей мутацию: спонтанные (причина не ясна) и индуцированные (вызываются мутагенами).
Мутагены – факторы среды, вызывающие мутацию. По природе мутагены бывают:
- физические (рентгеновское излучение);
- химические (асбест, формалин);
- биологические (ДНК вирусов).
Геномные мутации – связаны с изменением числа хромосом в геноме – гаплоидном наборе хромосом.
Различают 2 вида:
1. Полиплоидия (эуплоидия) — изменение числа хромосом, кратное гаплоидному набору. Различают автополиплоидию и аллополиплоидию.
а) При автополиплоидии многократно повторяется один и тот же хромосомный набор. Например, существуют виды хризантем, содержащие 18 хромосом (2n), 27 хромосом (3n), 36 хромосом (4n) … 81 хромосому (9n).
б) при аллополиплоидии многократно повторяются разные хромосомные наборы. Например, пшеница мягкая — гексаплоид, который содержит хромосомные наборы от 6 разных видов пшениц. Аллоплоидия в природе встречается редко, возникает при межвидовой гибридизации, чтобы гибриды смогли дать потомство необходима полиплоидизация. Так, селекционером Г.Д. Карпеченко был получен плодовитый гибрид капусты и редьки.
2. Гетероплоидия (анэуплоидия) — изменение числа хромосом, некратное гаплоидному набору. В результате возникают явления:
- трисомии (2n+1) — трисомия по 21 паре хромосом — синдром Дауна
- моносомии (2n-1) – моносомия по половым хромосомам – синдром Шерешевского-Тернера;
- нулесомии (2n-2).
Гетероплоидия обычно сопровождается серьезными наследственными аномалиями, часто несовместимыми с жизнью.
Хромосомные мутации - изменения структуры хромосом. Различают внутри- и межхромосомные аберрации (приведена классификация АА. Сазанова).
Внутрихромосомные перестройки:
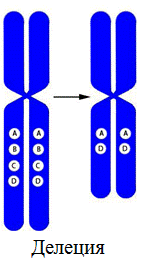
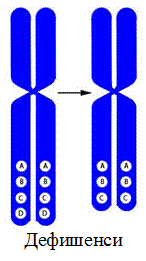
1. Делеция – утрата участка хромосомы.
2. Дефишенси – концевая делеция.
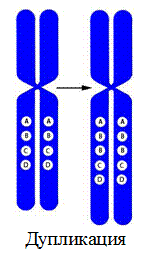
3. Дупликация – удвоение участка хромосомы.
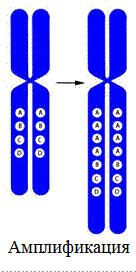
4. Амплификация – многократное повторение участка хромосомы.
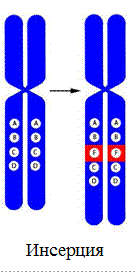
5. Инсерция – вставка дополнительного хромосомного района.
6. Парацентрическая инверсия – поворот на 180° участка хромосомы, не содержащего центромеру;
7. Перицентрическая инверсия – поворот на 180° участка хромосомы, содержащего центромеру.
|
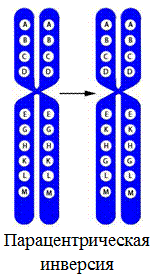
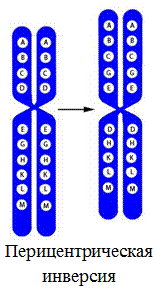
Межхромосомные перестройки:
1. Транслокация – перенос участка с одной хромосомы на другую;
2. Реципрокная транслокация – обмен участками между негомологичными хромосомами;
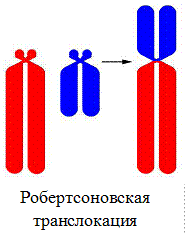
3. Робертсоновская транслокация – слияние двух акроцентрических хромосом с образованием одной субметацентрической хромосомы.
|
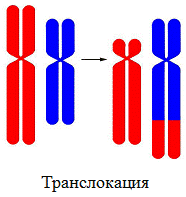
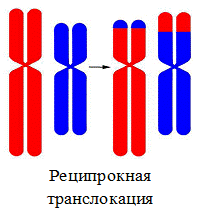
Генные мутации — связаны с изменением последовательности нуклеотидов в гене.
Различают 2 механизма:
1. Связан с изменением числа нуклеотидов (дупликации, делеции). В связи с тем, что генетический код не имеет знаков препинания, меняется состав всех кодирующих триплетов после места мутации, что приводит к замене многих аминокислот и синтезу белка с совершенно другими свойствами, что может оказаться губительно для организма.
2. Связан с заменой одного нуклеотида на другой. При этом меняется состав только одного триплета, что может привести к изменению только одной аминокислоты в белке (и то не всегда, так как генетический код вырожден). Замена 1 аминокислоты не всегда существенно сказывается на изменении свойств белка (особенно, если она по свойствам близка к исходной).
Генные мутации играют большую роль для эволюции. Они, по мнению С.С. Четверикова, создают резерв наследственной изменчивости. Так как большинство мутаций рецессивны, то в гетерозиготном состоянии могут долгое время себя не проявлять. Это очень важно, так как при изменении условий среды, выщепившаяся мутация может оказаться полезной и спасти вид от вымирания.
Генетика популяций
Отрасль генетики, изучающая генофонд популяции. Для изучения встречаемости отдельных генов и генотипов в популяции используют закон Харди-Вайнберга.
Закон Харди-Вайнберга был создан для идеальной популяции.
Черты идеальной популяции:
1. Популяция, характеризующаяся панмиксией – ничем не ограниченным свободным скрещиванием особей.
2. Популяция, на которую не действуют факторы эволюции, такие как естественный отбор, популяционные волны, дрейф генов.
3. Популяция, имеющая огромную численность.
Однако было установлено, что закон может быть с определенной долей условности применим для расчета доли определенных генов и генотипов и в реальной популяции.
Пусть p – частота гена А, тогда q – частота гена а.
p+q=1
(p(А)+q(а))2= p2(АА)+2 pq(Аа) +q2(аа)
Пример задачи:
Скрестили 60% особей с генотипом АА и 40% особей с генотипом аа. Определите вероятность генотипов АА, Аа и аа после установления в популяции динамического равновесия.
Из условия задачи делаем вывод, что p=0,6, а q=0,4. Тогда
Частота генотипа АА (p2 )= 0,62=0,36
Частота генотипа Аа (2 pq)= 2х0,6х0,4=0,48
Частота генотипа аа (q2)=0,42=0,16
Интересные вопросы по генетике
1. В каких структурно-функциональных состояниях и где (в каких организмах, структурах) находится функционирующая или сохраняющая способность к функционированию ДНК?
Функционирующая или сохраняющая способность к функционированию («живая») ДНК может находиться:
· в составе эухроматина клеточного ядра в интерфазе жизненного цикла клетки; ДНК, образующая эухроматин, является активной, участвующей в транскрипции;
· в составе гетерохроматина, который, в отличие от локализованного в центре ядра эухроматина, располагается по периферии клеточного ядра; образующая гетерохроматин ДНК функционально неактивна, т.е. не участвует в транскрипции;
· в виде кольцевой молекулы ДНК клеточных органоидов – митохондрий и пластид; эта ДНК, кодирующая преимущественно структурные белки органоидов, обычно является всегда активной;
· в виде кольцевой молекулы ДНК прокариотической клетки; локализованная непосредственно в цитоплазме, эта ДНК функционирует непрерывно;
· в составе хроматид (дочерних хромосом) в период митоза: отличаясь высшей степенью спирализации и связи с упаковочными белками-гистонами, эта ДНК является неактивной;
· в составе многих вирусов, образуя их наследственный аппарат, окруженный белковым чехлом (капсидом); вирусная ДНК переходит в активное состояние только внутри клетки организма – хозяина;
· в виде плазмид, которые устроены еще проще, чем вирусы: представляют собой лишенные какого-либо окружения («обнаженные») молекулы или фрагменты молекул ДНК; они локализуются в клетке бактерий, где могут либо соединяться с кольцевой молекулой ДНК, либо оставаться непосредственно в цитоплазме; плазмидная ДНК является активно функционирующей ДНК, придающей прокариотической клетке дополнительные наследственные свойства: например, пеницилиназная плазмида (R-фактор или фактор устойчивости) обеспечивает устойчивость стафилококков к пенициллину.
.
2. Какой район рассматривался академиком Н.И.Вавиловым в качестве «центра происхождения культурных растений»?
В качестве «центра происхождения культурных растений» Н.И.Вавиловым рассматривался район, в котором обнаруживалось наибольшее генетическое разнообразие представителей данного вида растений. Н.И.Вавилов выделил 7 центров происхождения культурных растений:
· южно-азиатский (рис, цитрусовые, сахарный тростник);
· восточно-азиатский или китайский (соя, просо, гречиха);
· юго-западноазиатский (пшеница, рожь, бобовые, виноград, плодовые культуры);
· средиземноморский (капуста, маслины);
· абиссинский (ячмень, кофейное дерево);
· центральноамериканский (хлопок, кукуруза, фасоль, какао);
· южноамериканский (картофель, хинное дерево).
В качестве древнейшего среди них он рассматривал абиссинский центр. Впоследствии число центров происхождения культурных растений было увеличено до 12.
Дата добавления: 2018-11-25; просмотров: 1575;