Пластичность поведения простейших
Как мы видим, и в моторной и в сенсорной сфере поведение достигает у ряда видов простейших известной сложности. Достаточно указать на фобическую реакцию (реакцию испуга) туфельки в вышеописанном примере клинотаксиса: наткнувшись на твердое препятствие (или попав в иную неблагоприятную зону), туфелька резко останавливается и принимает «оборонительное положение», т.е. «съеживается», готовясь пустить в ход ядовитые стрекательные капсулы. Одновременно меняютсядвижения ресничек, происходит тактильное и химическое обследование объекта и т.д. У эвглены фобическая реакция выражается в том, что она, остановившись, начинает производить передним концом тела круговые движения, после чего уплывает в другом направлении (рис. 29).
Ясно, что такая интеграция моторно-сенсорной активности возможна лишь с помощью специальных функциональных структур, аналогичных нервной системе многоклеточных животных. Однако о морфологии этих аналогов еще очень мало известно, и только относительно инфузории удалось с определенной достоверностью доказать существование специальной сетевидной системы проводящих путей, располагающейся в эктоплазме. Очевидно, проведение импульсов осуществляется у простейших и системой градиентов в самой цитоплазме.
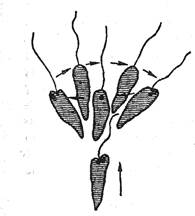
Рис. 29. Фобическая реакция эвглены при внезапном изменении освещения (по Джен-нингсу)
Способность к индивидуальному изменению описанных генетически фиксированных форм поведения путем научения выражена у простейших слабо. Все же способность простейших к научению, хотя бы в элементарных формах, сейчас можно считать доказанной. Если, например, держать парамеции некоторое время в треугольном или квадратном (в сечении) сосуде, то они сохраняют привычный путь передвижения вдоль стенок сосуда и после их перемещения в сосуд округлой формы (рис. 30). Аналогичным образом инфузория, плававшая длительное время (около двух часов) в сосуде, имевшем форму треугольника, придерживается этой траектории затем и в квадратном сосуде большей площади (опыты немецкого ученого Ф. Брамштедта). В опытах советской исследовательницыН.А. Тушмаловой инфузории были подвергнуты постоянно действующему ритмичному раздражителю — вибрации. В результате животные постепенно переставали реагировать на него обычным образом (сокращением). Исследовательница усматривает в этом пример элементарных следовых реакций, представляющих собой простую форму кратковременной памяти, формирующейся на этом уровне филогенеза за счет чисто молекулярных взаимодействий Подобные опыты ставились и другими экспериментаторами, применявшими различные формы воздействия. Высказывались сомнения, действительно ли в таких случаях можно говорить о каких-то формах научения, ибо сильные воздействия могли оказать хотя бы временное вредное влияние на подопытных животных. К тому же вообще в опытах с простейшими очень трудно учесть все возможные побочные факторы (особенно химические изменения среды).

Рис. 30. Привыкание у парамеции (по Брамштедту). Пояснения см. в тексте
Все же, очевидно, мы имеем здесь дело с элементарной формой научения — привыканием. Привыкание (к измененным внешним условиям), как мы уже знаем, играет немаловажную роль и в поведении высших животных, но имеет качественно иной характер уже хотя бы потому, что строится у них не только на ощущениях, но и на восприятиях. При элементарной же сенсорной психике животное может привыкать лишь к воздействию отдельных раздражителей (или их совокупностей), являющихся воплощением отдельных свойств или качеств компонентов окружающей среды. Именно это и имело место в приведенных примерах, когда наблюдалась модификация ви-дотипичного поведения: врожденные реакции на определенные раздражители постепенно элиминируются, если при многократном повторении не наступит биологически значимый эффект. Так, в опытах Тушмаловой фобическая реакция у инфузорий со временем сводилась на нет при повторной встрече с «безобидным» раздражителем.
Привыкание необходимо отличать от утомления, выражающегося в явлениях «истощения» животного. В опытах Тушмаловой это выражалось в неспособности инфузории к дальнейшему реагированию, если она в течение 2—3 часов подвергалась очень сильным раздражениям. Утомление связано с перерасходом энергетических ресурсов, привыкание же представляет собой активную приспособительную реакцию, значение которой состоит в экономии этих ресурсов, в предотвращении траты энергии на бесполезные для животного движения.
Как форма научения привыкание характеризует низший уровень элементарной сенсорной психики, хотя и не теряет своего значения на всех ступенях развития психики, как это было показано при ознакомлении с онтогенезом поведения животных (см. ч. II). Однако у высших представителей простейших, возможно, уже существуют и зачатки ассоциативного научения, вообще характерного для более высоких уровней психического развития.
Об ассоциативном научении можно говорить в тех случаях, когда устанавливается временная связь между биологически значимым и «нейтральным», точнее, биологически маловалентным раздражителем. Именно такую связь Брамштедту удалось выработать у туфельки, которая, как уже отмечалось, не реагирует заметным образом на изменение освещения, но весьма чувствительна к температурным изменениям. Если в эксперименте затенить одну половину капли воды, в которой плывут парамеции, и при этом подогреть освещенную часть капли, то вскоре инфузории соберутся в холодной темной части, но останутся там (в течение 15 минут) и после того, как температура в обеих частях капли будет уравновешена. Правда, эти опыты Брамштедта подвергались серьезной критике, так как при подогреве меняется и химизм воды (меняется растворимость содержащихся в ней газов), что не может не влиять на поведение инфузории (опыты У.Грабовского).
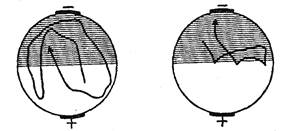
Рис. 31. Образование у парамеции временной связи между электрическим раздражением и светом (по Зесту). Пояснения см. в тексте
Вместе с тем аналогичный результат может быть получен и путем наказания животных при заплыве в освещенную зону ударами электрического тока (рис. 31). Эффект избегания сохранялся в этом случае в течение 20 минут (опыты Г. Зеста). Однако обратный результат, т.е. избегание темноты и заплыв в освещенную зону, Зесту получить не удалось. Это обстоятельство, однако, является результатом методической ошибки: инфузориям свойствен гальванотаксис, выражающийся в их устремлении к катоду. В упомянутых же опытах Зеста катод всегда находился на темной половине и гальванотаксис оказывался сильнее электрического раздражения, почему инфузории и направлялись только в темноту.
В более тщательно поставленных опытах польскому ученому С. Вавржинчику удалось преодолеть эти методические недостатки и с успехом научить инфузории избегать темный участок стеклянной трубки, где их раздражали электрическим током. Постепенно туфельки все чаще оставались в освещенной зоне и поворачивали на границе с затемненной зоной даже при перемещении этой границы вдоль трубки еще до получения электрического удара. Наконец, они оставались в освещенной зоне в четыре разадольше, чем в темной, даже тогда, когда раздражения током совсем прекратились. Эта реакция сохранялась даже в течение 50 минут, что является достаточно большим сроком для простейших. Четкие положительные результаты этот исследователь получил и в других экспериментах.
Правда, Я. Дембовский, повторивший эти опыты с некоторыми изменениями, пришел к выводу, что, хотя и не исключена возможность выработки у простейших обусловленных реакций, эти опыты с туфельками все-таки не решают до конца сути вопроса, ибо существуют несравненно более сильные факторы, особенно химические, которые маскируют реакции на свет. Ввиду отсутствия вполне убедительных экспериментальных данных Дембовский оставил вопрос о наличии у простейших обусловленных реакций открытым.
Вместе с тем и у других (кроме туфельки) инфузорий, в частности у стентора и сувойки, удалось после 140—160 сочетаний выработать сократительные реакции в ответ на световое раздражение.
На инфузориях проводились и другие опыты с применением разных методических приемов. В частности, использовались капилляры, иногда с загнутым концом, в которые сажались парамеции, после чего измерялось время, необходимое им для высвобождения. В новейших исследованиях, проведенных таким способом, получались вполне положительные результаты: при каждом повторении опыта время выхода парамеции из трубочки заметно уменьшалось, что и признается доказательством их способности к ассоциативному научению.
Однако американские ученые Ф.Б. Эпплуайт и Ф.Т. Гарднер, недавно повторившие эти опыты, считают такой вывод необоснованным. Эти ученые засасывали последовательно в один и тот же капилляр разные, еще ни разу не побывавшие в опыте туфельки и установили, что время выхода этих животных из капилляра также постепенно уменьшается. Но этого не случалось, если капилляр до употребления тщательно промывался и кипятился в деионизированной воде. Использование обработанных такимобразом капилляров для повторного проведения опыта с одной и той же туфелькой не давало практически никакого эффекта научения. Отсюда экспериментаторы вывели заключение, что уменьшение времени выхода из капилляра является у парамеции не результатом научения, а реакцией на загрязнение внутренней поверхности трубки продуктами обмена веществ, которое увеличивается от опыта к опыту и от которого животные пытаются все скорее уйти.
С другой стороны, в аналогичных исследованиях других ученых учитывались подобные возможные изменения среды, и было установлено, что время выхода инфузорий из трубки сокращается лишь в первых опытах, потом же оно остается постоянным (среда в трубке должна, однако, продолжать изменяться).
Таким образом, вопрос о наличии у инфузорий (а тем более у простейших вообще) ассоциативного научения еще нельзя считать решенным. Все же, очевидно, такое научение существует у них в зачаточном виде.
Дата добавления: 2017-02-20; просмотров: 435;