Сравнительный обзор развития двигательной активности зародышей
Беспозвоночные
Эмбриональное поведение беспозвоночных еще очень слабо изучено. Те немногие сведения, которые пока получены, относятся в основном к кольчатым червям, моллюскам и членистоногим. Известно, например, что зародыши головоногих моллюсков уже на ранних стадиях своего развития вращаются внутри яйца вокруг своей оси со скоростью, один оборот в час. В других случаях зародыши передвигаются от одного полюса яйца к другому. Все эти движения осуществляются с помощью ресничек, что представляет особый интерес, если учесть, что этот примитивный способ передвижения у взрослых головоногих моллюсков отсутствует, но широко распространен среди личинок морских беспозвоночных.
Заслуживает также внимания, что к концу эмбриогенеза у беспозвоночных оказываются уже вполне сформированными некоторые инстинктивные реакции, имеющие первостепенное значение для выживания. У мизид (морские ракообразные), например, к моменту вылупления уже вполне развита реакция избегания, т.е. уклонения от неблагоприятных воздействий. Но, как показал канадский исследователь М. Беррилл, первоначально у эмбриона наблюдается спонтанное «вздрагивание» и «подергивание», которые затем все чаще возникают в ответ на прикосновение к икринке. Таким образом, первоначально спонтанные движения постепенно сменяются рефлекторными.
У других ракообразных — морских козочек (отряд бокоплавы) — с 11-го по 14-й день развития, т.е. до вылупления, наблюдаются спонтанные и ритмичные движения головы и других частей эмбриона, из которых впоследствии формируются специфические двигательные реакции этих рачков. Только к концу эмбриогенеза, в день вылупления, появляются двигательные ответы на тактильные раздражения (прикосновения волоском к зародышу, с которого в эксперименте была снята яйцевая оболочка). Вестественных условиях весь набор движений взрослой особи обнаруживается уже спустя 10 часов после вылупления.
У эмбриона дафнии антенны, служащие у взрослых особей для плавания, начинают двигаться на средних этапах эмбриогенеза, а перед его завершением они поднимаются и принимают то положение, которое необходимо для выполнения плавательных движений. В это время (за несколько часов до выхода из яйца) антенны начинают особенно интенсивно двигаться.
В этих примерах отчетливо проявляется постепенное формирование рефлекторного ответа на основе первоначально эндогенно обусловленных движений, которые впоследствии связываются с внешними раздражителями, отчасти уже путем «эмбрионального научения». Этот процесс сопряжен с глубокими морфологическими преобразованиями.
Низшие позвоночные
Первые движения зародышей рыб, по данным ряда исследователей, также возникают спонтанно на эндогенной основе. Еще в 20-х годах было показано, что движения зачатков органов появляются в строгой последовательности, в зависимости от созревания соответствующих нервных связей. После появления сенсорных нервных элементов на поведение зародыша начинают влиять и экзогенные факторы (например, прикосновения), которые сочетаются с генетически предопределенной координацией движений. Постепенно первоначально генерализованные движения зародыша дифференцируются.
Вообще у зародышей костистых рыб обнаруживаются к концу эмбриогенеза такие движения: дрожание, подергивание отдельных частей тела, вращение, змеевидное изгибание. Кроме того, перед вылуплением производятся своеобразные «клевательные» движения, облегчающие выход из яйцевидной оболочки. Кроме того, выклеву способствуют и изгибательные движения тела. В ряде случаев удалось установить четкую связь между появлением новых двигательных актов и общим анатомическим развитием.
Сходным образом совершается формирование эмбрионального поведения и у земноводных. Из первоначально генерализованного сгибания всего тела зародыша постепенно формируются плавательные движения, движения конечностей и т.д., причем и здесь двигательная активность развивается первично на эндогенной основе.
Интересный пример представляет в этом отношении жаба Eleutherodactylus martinicensis с острова Ямайка, у которой выход из икринки как бы задерживается и личинка развивается внутри яйцевых оболочек. Тем не менее у нее проявляются все движения, свойственные свободноплавающим личинкам (головастикам) других бесхвостых земноводных. Как и у последних, плавательные движения формируются у этой личинки постепенно из более генерализованных двигательных компонентов: первые движения конечностей еще слиты с общим извиванием всего тела, но уже спустя сутки можно вызвать одиночные рефлекторные движения одних конечностей независимо от движений мышц туловища; несколько позже и в строгой последовательности появляются более дифференцированные и согласованные движения всех четырех конечностей, и наконец возникают во всех деталях вполне координированные плавательные движения с участием всех соответствующих моторных компонентов, хотя плавать сформировавшаяся к этому времени личинка еще не начинала, ибо она по-прежнему заключена в яйцевые оболочки.
Что же касается хвостатых амфибий, то Когхилл показал в свое время, что эмбрион амбистомы производит плавательные движения еще задолго до вылупления, изгибаясь сперва наподобие буквы «С» и позже как буква «S». Еще позже появляются движения ног, типичные для передвижения по суше взрослой амбистомы, причем нейромышечная система, детерминирующая первичные плавательные движения, определяет и эти локомоторные элементы, особенно последовательность и ритм движений. Кармайкл сумел на этом же объекте доказать, что этот механизм созревает без научения. Он вырастил эмбриона амбистомыв анестезирующем растворе ацетонхлороформа, который полностью обездвиживал зародыш, но не препятствовал его росту и морфогенезу. В таких условиях локомоторные способности развивались вполне нормально, и в итоге они не отличались от таковых контрольных организмов, развитие которых совершалось в обычных условиях. Из своих опытов Кармайкл вывел заключение, что формирование способности к плаванию не нуждается в процессах научения, а зависит исключительно от анатомического развития.
Однако, как справедливо отмечал по поводу этих опытов польский зоопсихолог Я. Дембовский, у подопытных эмбрионов подавлялись лишь мышечные движения, возможность накопления двигательного эмбрионального опыта, но не другие функции, в частности процессы в развивающейся нервной системе. Невозможно, как пишет Дембовский, создать такие экспериментальные условия, при которых нервная система развивалась бы, не функционируя. Поскольку нервная система начинает функционировать еще до того, как она окончательно сформировалась, это функционирование также является своего рода упражнением, которое в свою очередь является важным фактором развития нервной деятельности, а тем самым всего поведения зародыша.
Для выявления эндогенной обусловленности формирования двигательной активности зародышей производились интересные опыты на эмбрионах саламандр: пересаживались зачатки конечностей таким образом, что последние оказывались повернутыми в обратную сторону. Если бы формирование их движений определялось эмбриональным упражнением (путем афферентной обратной связи), то в ходе эмбриогенеза должна была бы произойти соответствующая функциональная коррекция, восстанавливающая способности к нормальному поступательному движению. Однако этого не произошло, и после вылупления животные с повернутыми назад конечностями пятились от раздражителей, которые у нормальных особей обусловливают движение вперед. Сходные результаты былиполучены и у эмбрионов лягушек: перевертывание зачатков глазных яблок на 180° привело к тому, что оптокинетические реакции оказались у этих животных смещенными в обратном направлении (рис. 19).
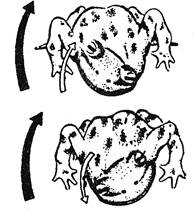
Рис. 19. Инверсии оптокинетической реакции у лягушек (опыт Сперри). Большие черные стрелки обозначают направление оптического раздражения, вызывающего оптокинетическую реакцию (маленькая светлая стрелка) в норме (вверху) и после поворота зачатков глазных яблок на 180° (внизу)
Все эти данные приводят к заключению, что формирование в эмбриогенезе локомоторных движений и оптомоторных реакций (а также некоторых других проявлений двигательной активности) происходит у низших позвоночных, очевидно, не под решающим влиянием экзогенных факторов, а в результате эндогенно обусловленного созревания внутренних функциональных структур.
Птицы
Эмбриональное поведение птиц изучалось преимущественно на зародышах домашней курицы. Уже в конце вторых суток появляется сердцебиение, а начало спонтанной двигательной активности куриного эмбриона приурочено к 4-му дню инкубации. Весь период инкубации длится три недели. Движения начинаются с головного конца и постепенно простираются к заднему, охватывая все большие участки тела зародыша. Самостоятельные движения органов (конечностей, хвоста, головы, клюва, глазных яблок) появляются позже.
Как уже говорилось, Куо установил наличие и показал значение эмбриональной тренировки у зародыша курицы (а также других птиц), но впал при этом в односторонность, отрицая наличие врожденных компонентов поведения и спонтанной активности как таковой. В дополнение к сказанному приведем еще несколько примеров из работ этого выдающегося исследователя.
Куо установил, что максимальная двигательная активность эмбриона совпадает по времени с движениями амниона, т.е. внутренней зародышевой оболочки, обволакивающей зародыш. Из этого Куо заключил, что пульсация оболочки обусловливает начато двигательной активности последнего. Впоследствии, однако, В. Гамбургером было показано, что нет подлинной синхронизации между этими двигательными ритмами, а Р.В.Оппенгейм экспериментально доказал, что движения эмбриона не только не зависят от движения амниона, а, возможно, даже сами обусловливают их.
Формирование клевательных движений, по Куо, первично определяется ритмом биения сердца зародыша, ибо первые движения клюва, его открывание и закрывание, совершаются синхронно с сокращениями сердца. Впоследствии эти движения коррелируются со сгибательными движениями шеи, а незадолго до вылупления клевательный акт следует за любым раздражением тела в любой его части. Таким образом, реакция клевания, сформировавшаяся путем эмбриональной тренировки, имеет к моменту вылупления птенца весьма генерализованный характер. «Сужение» реакции в ответ на действие лишь биологически адекватных раздражителей происходит на первых этапах постэмбрионального развития. По Куо, так обстоит дело и с другими реакциями.
Важную роль в формировании эмбрионального поведения Куо отводил также непосредственному влиянию среды, окружающей зародыш, и происходящим в ней изменениям. Так, например, у куриного зародыша начиная с 11-го дня инкубации желточный мешок начинает надвигаться на брюшную сторону зародыша, все больше стесняя движения ног, которые до вылупления сохраняют согнутое положение, причем одна нога располагается поверх другой. По мере того как желток всасывается зародышем, верхняя нога получает все большую свободу движения, однако находящаяся под ней нога по-прежнему лишена такой возможности. Лишь после того как верхняя нога получит возможность достаточно далеко отодвигаться, нижняя может начать двигаться.
Таким образом, движения ног развиваются с самого начала не одновременно, а последовательно. Куо считал, что именно этим определяется развитие механизма последовательных движений ног цыпленка при ходьбе, что здесь кроется первопричина того, что вылупившийся из яйца цыпленок передвигается шагами, попеременно переставляя ноги, а не прыжками, отталкиваясь одновременно обеими лапками. Поддерживая концепцию Куо, Боровский писал по этому поводу, что «цыпленок шагает не потому, что у него имеется "инстинкт шагания", а потому, что иначе не могут работать его ноги и их механизм, выросшие и развившиеся в таких именно условиях. Механизм этот не появится как нечто готовое в тот момент, когда он понадобится, а развивается определенным путем, под влиянием других факторов»3.
Многие механистические концепции Куо не выдержали экспериментальной проверки, произведенной более поздними исследователями, другие еще нуждаются в такой проверке, тем более что вскрытые им коррелятивные отношения не обязательно указывают на причинные связи. Не подтвердилось, в частности, мнение Куо о том, что ведущим фактором двигательной активности зародыша в раннем эмбриогенезе является сердцебиение.
Гамбургером и его сотрудниками было установлено., что уже на ранних стадиях эмбриогенеза движения зародыша имеют нейрогенное происхождение. Электрофизиологические исследования показали, что уже первые движения обусловливаются спонтанными эндогенными процессами в нервных структурах куриного эмбриона. Спустя 3,5—4 дня после появления первых его движений наблюдались первые экстероцептивные рефлексы, однако Гамбургер, Оппенгейм и другие показали, что тактильная, точнее, тактильно-проприоцептивная стимуляция не оказывает существенного влияния на частоту и периодичность движений, производимых куриным эмбрионом на протяжении первых 2—2,5 недель инкубации. По Гамбургеру, двигательная активность зародыша на начальных этапах эмбриогенеза «самогенерируется» в центральной нервной системе.
Гамбургером производился следующий опыт: перерезав зачаток спинного мозга в первый же день развития куриного эмбриона, он регистрировал впоследствии (на 7-й день эмбриогенеза) ритмичные движения зачатков передних и задних конечностей. Нормально эти движения протекают синхронно. У оперированных же эмбрионов эта согласованность нарушилась, но сохранилась самостоятельная ритмичность движений.
Это указывает на независимое эндогенное происхождение этих движений, а тем самым и соответствующих нервных импульсов, на автономную активность процессов в отдельных участках спинного мозга. С развитием головного мозга он начинает контролировать эти ритмы. Вместе с тем эти данные свидетельствуют о том, что двигательная активность не обусловливается исключительно обменом веществ, например, такими факторами, как уровни накопления продуктов обмена веществ или снабжения тканей кислородом, как это принималось некоторыми учеными.
При изучении эмбрионального развития поведения птиц необходимо учитывать специфические особенности биологии исследуемого вида, которые отражаются и на протекании эмбриогенеза. Особенно это касается различий между выводковыми (зрелорождающимися) и птенцовыми (незрелорождающимися) птицами. Так, например, как показал советский исследователь Д.Н.Гофман, по сравнению с курицей грач развивается более ускоренно, быстрее накапливается масса тела зародыша, зато у курицы эмбриогенез проходит более равномерно и имеется больше периодов роста и дифференциации. Последний период формирования морфологических структур и поведения проходит у курицы еще внутри яйца, у грача же (как не-зрелорождающейся птицы) этот период относится к постэмбриональному развитию.
Млекопитающие
В отличие от рассмотренных до сих пор животных зародыши млекопитающих развиваются в утробе матери, что существенно усложняет (и без того весьма трудное) изучение их поведения, поэтому по эмбриональному поведению млекопитающих накоплено значительно меньше данных, чем по куриному эмбриону и зародышам земноводных и рыб. Непосредственные визуальные наблюдения возможны лишь на эмбрионах, извлеченных из материнского организма, что резко искажает нормальные условия их жизни. Рентгенологические исследования указывают на то, что двигательная активность таких искусственно изолированных зародышей выше, чем в норме. Вместе с тем именно на таких объектах, преимущественно зародышах грызунов, были получены те данные, которыми мы сегодня располагаем.
Так, например, по Кармайклу, развитие двигательной активности совершается у зародыша морской свинки следующим образом. Первые движения состоят в подергивании шейно-плечевого участка туловища эмбриона. Они появляются приблизительно на 28-й день после оплодотворения. Постепенно появляются и другие весьма разнообразные движения, а к 53-му дню, т.е. приблизительно за неделю до родов, формируются четко выраженные реакции, которые достигают максимального развития за несколько дней до рождения. У такого эмбриона обнаруживаются уже вполне адекватные, и главное, видоизменяющиеся рефлекторные ответы на тактильные раздражения: прикосновение волоском к коже около уха вызывает специфическое подергивание последнего, непрерывное продолжение этого раздражения или его многократное повторение — приведение конечности данной стороны к раздражителю, чем нередко достигается его удаление; еслиже и после этого продолжать раздражение, то приходит в движение вся голова, а затем и туловище эмбриона, что может привести к его вращению, и наконец, по выражению Кармайкла, «каждая мышца приводится в действие». Кармайкл говорил по этому поводу, что эмбрион ведет себя как бы согласно пословице: «если не сразу достиг успеха — пробуй, пробуй еще», но подчеркивал, что нет оснований предполагать, чтобы что-либо в этом поведении являлось выученным.
Эмбриональное развитие поведения млекопитающих существенно отличается от такового у других животных. Это отличие выражается в том, что у млекопитающих движения конечностей формируются не из первоначальных общих движений всего зародыша, как мы это видели у вышеупомянутых других позвоночных, особенно низших, а появляются одновременно с этими движениями или даже раньше их. Вероятно, в эмбриогенезе млекопитающих большее значение приобрела ранняя афферентация, чем спонтанная эндогенная нейростимуляция.
Постоянная тесная связь развивающегося зародыша с материнским организмом, в частности посредством специального органа — плаценты, создает у млекопитающих совершенно особые условия для развития эмбрионального поведения. Новым и весьма важным фактором является в этом отношении возможность воздействия на этот процесс со стороны материнского организма, прежде всего гуморальным путем.
На такую возможность косвенно указывают результаты экспериментов, при которых на женских зародышей морской свинки еще во время их внутриутробного развития воздействовали мужским половым гормоном (тестостероном). В результате, став половозрелыми, они проявили признаки самцового поведения в ущерб сексуальному поведению, свойственному нормальным самкам. Аналогичное воздействие, произведенное после рождения, не давало такого эффекта. Подобным образом удавалось преобразовывать и половое поведение мужских особей. Очевидно, в ходе эмбриогенеза содержание тестостерона ворганизме зародыша влияет на формирование центральных нервных структур, регулирующих сексуальное поведение: его отсутствие — в сторону женских признаков, его наличие — в сторону мужских.
В экспериментах ряда исследований у беременных самок крыс периодически вызывали состояния беспокойства. В таких условиях родились более пугливые и возбудимые детеныши, чем в норме, несмотря на то, что их затем вскармливали другие самки, не подвергавшиеся экспериментальным воздействиям. Эти данные особенно отчетливо показывают роль влияния материнского организма на формирование признаков поведения детеныша в эмбриональном периоде его развития.
Дата добавления: 2017-02-20; просмотров: 638;