Сравнительный анализ фаун
Изучение структуры фауны является первым этапом ее исследования. Но этого недостаточно, чтобы понять ее особенности и в дальнейшем судить о ее происхождении. Специфика одной фауны, ее отличия от других становятся очевидными только при сравнении их структур. Для этого обычно берутся две соседние фауны, в отдельных же случаях требуется сравнение и отдаленных фаун. Необходимым условием такого рода исследований является примерно равная степень изученности сравниваемых фаун, хотя это не всегда соблюдается из-за неполноты фаунистических сведений.
Сравнительный анализ начинается с изучения списков видов локальных фаун. При этом обращается внимание на качественные отличия, т.е. на таксоны (виды, роды и т.д.), которые присутствуют лишь в одной из сравниваемых фаун. Количество видов (или других таксонов), характерных только для одной локальной фауны, служит показателем ее специфики.
При сравнительном анализе большое значение имеют определение степени общности и различия между двумя фаунами. Более точно это можно установить с помощью количественных методов. Для математического выражения степени сходства фаун двух районов предложен ряд формул. Наиболее часто применяется формула Жаккара, по которой определяют коэффициент общности (К), выражающийся в процентах:
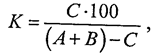
где С — число видов, общих для обеих фаун;
А, В — число видов соответственно в первой и второй фауне.
Используется также формула Сьеренсена:
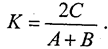
Оба коэффициента в общем однотипно отражают степень сходства фаун сравниваемых районов. Применяют и другие показатели, например индекс различия фаун.
Эндемизм
Одной из важнейших особенностей фауны является эндемизм. Эндемичными систематическими единицами называются виды, роды и другие таксономические единицы, которые встречаются только в какой-то определенной области и нигде более. В силу этого эндемики представляют специфический компонент любой фауны и абсолютно отличают ее от других фаун. Если более широко распространенные виды указывают на связи между фаунами, то эндемики резко отличают их друг от друга.
Количество эндемиков в различных фаунах неодинаково. Наиболее высок процент эндемизма в островных фаунах, а на континентах — в районах с сильно расчлененным рельефом, т. е. в горных странах, поскольку географическая изоляция — необходимое условие процесса видообразования.
Для образования эндемичных видов требуется не только географическая изоляция, но и продолжительное время. Длительное и непрерывное развитие фауны благоприятствует возникновению новых видов, в то время как быстрые изменения условий существования, наоборот, ведут к вымиранию одних и расселению других. Обилие эндемиков отражает длительность развития фаун.
Различия между эндемичными и неэндемичными компонентами фауны не абсолютны. В большинстве фаун есть так называемые субэндемики, или почти эндемичные таксоны (роды, виды).
В историческом смысле можно говорить о прогрессивном и реликтовом эндемизме. Прогрессивными эндемиками (неоэндеми-ками) считаются виды или роды, возникшие в фауне недавно и еще не распространившиеся за пределы ее территории. Это молодые новообразования, которые незначительно отличаются от остальных видов этого же рода. Между ними и другими видами, обитающими на одной территории, существуют легко различимые связи. Часто прогрессивные эндемики образуют группы близкородственных видов. Их экологические требования соответствуют окружающим условиям среды.
К реликтовым эндемикам (палеоэндемикам) относятся животные, ареал которых в прошлом был гораздо больше, а в настоящее время они сохранились лишь на отдельной территории. Реликтовые эндемики (их иногда называют консервативными) занимают в фауне, как правило, изолированное положение. Близкородственных видов у них нет, они систематически обособлены. Реликтовые эндемичные роды часто включают лишь несколько или всего-навсего один вид (монотипичные роды). Экологическая обстановка их «устраивает» относительно. Конечно, если бы она полностью не отвечала их потребностям, они бы давно вымерли. Однако распространение этих видов в ареале носит пятнистый характер, они узколокальны и связаны с ограниченными биотопами. Наличие в фауне реликтовых эндемиков говорит о том, что благоприятная для них обстановка сохраняется длительное (в геологическом смысле) время, пусть даже на небольшом пространстве. Замечательным примером реликтового эндемизма служит гаттерия Sphenodon punctatus — единственный представитель семейства клинозубых Sphenodontidae, отряда клювоголовых Rhyn-chocephalia рептилий, живущая в Новой Зеландии. Для консервативных эндемиков высокого ранга нередко характерно дизъюнктивное распространение.
Генезис фуны
Каждая фауна имеет свою историю. Формирование фауны, или фауногенез, продолжается очень длительное время, измеряемое иногда целыми геологическими периодами. Те виды и группы животных, которые живут в настоящее время на той или иной территории, могут быть весьма отдаленными потомками других животных, давно вымерших либо больше не встречающихся в каких-то частях Земли. Описанием процесса зарождения и смены фаун занимается генетическая, или историческая, зоогеография. Для того чтобы ее выводы и заключения были достоверными, необходимо привлечь целый арсенал прямых и косвенных доказательств смены комплекса видов животных во времени и пространстве. Лучшую опору для познания прежнего облика фауны дают ископаемые находки. Они и являются прямыми доказательствами пребывания какого-нибудь вида в составе изучаемой фауны в определенный период ее развития. Примерами могут служить находки скелетов мамонта в Сибири или окаменевших костей динозавров в Монголии.
Но такие находки редки, а многие группы животных не сохраняются в ископаемом состоянии вообще. При отсутствии палеонтологических материалов характер фауногенеза устанавливается по косвенным свидетельствам — данным исторической геологии, палеогеографии, выводам на основе теории эволюции. Большую пользу приносят также и собственные зоогеографические сведения о дизъюнкциях ареалов, эндемиках и реликтах.
Островные фауны
Своеобразие фауны островов интересовало ученых с момента зарождения зоогеографии как науки. В ряде случаев изучение особенностей островной биоты приводило к появлению крупных обобщений и биологических теорий. Так, знакомство Ч. Дарвина с фауной Галапагосских островов навело его на мысль о дивергенции близких видов, происходящих от общего предка. Специально фауне островов А. Уоллес посвятил книгу «Островная жизнь» (1880). Острова интересуют биологов, во-первых, из-за относительной простоты структуры их биоценозов и трофических связей между видами, что облегчает изучение сообщества, а во-вторых, из-за необычных, порой удивительных форм жизни.
Все разнообразие островов Мирового океана можно свести к нескольким основным типам. Главным критерием выделения типов является происхождение островов. Чаще всего они бывают океаническими и материковыми. Океанические острова никогда не были частями материков, а образовались в результате поднятия дна океана, либо извержения подводных вулканов и застывания лавы, либо, наконец, вследствие жизнедеятельности рифообразующих кораллов. Примерами подобных островов являются архипелаги Микронезии и Полинезии в Тихом океане, остров Св. Елены и др.
Материковые, или континентальные, острова представляют собой части материков, отделившиеся от них в определенную геологическую эпоху. К ним относятся Британские, Большие Зонд-ские (Ява, Суматра, Калимантан и др.), Тайвань, Новая Гвинея, Тасмания и многие другие.
Остров материкового происхождения при отделении от материка наследует тот или иной (но не полный) набор местообитаний и связанных с ним видов. С одной стороны, этим определяется умеренное зоогеографическое своеобразие таких островов: с момента их отделения от материка начинается дифференцировка некоторых видов, образуются местные формы (подвиды, виды, реже роды). С другой стороны, происходит объединение первоначальной фауны за счет исчезновения ряда видов, причины которого будут рассмотрены ниже. Наконец, на материковых островах отсутствуют виды или роды, возникшие на материках или проникшие на них после отделения острова. Островная фауна в любом случае будет беднее предковой материковой.
На острове океанического происхождения первое время после его образования вообще нет никаких организмов. Современный органический мир таких островов формируется за счет видов, попавших на них путем залетов или заносов, т. е. активным или пассивным путем. В некоторых же случаях в фауну входят обитатели океана, приспособившиеся к жизни на суше: некоторые рыбы, раки (например, пальмовый вор), пресноводные моллюски и др. Фауна океанических островов от начала своего образования и до достижения устойчивого состояния непрерывно обогащается.
Независимо от происхождения фауна острова несет «островной» отпечаток, степень которого зависит от площади острова, расстояния его до ближайшей суши, от климатической зоны, в которой он располагается, и т.д. Иначе говоря, существуют конкретные признаки островных фаун, 4 из них являются основными: бедность и дефектность, выраженный эндемизм, ограниченное количество архаических форм и, наконец, адаптивная радиация в рамках небольшого количества эндемичных групп.
Относительная бедность и дефектность (систематическая неполнота) островных фаун объясняется тем, что площадь острова всегда меньше площади ближайшего материка и, стало быть, часть биотопов на нем отсутствует или занимает по сравнению с материком меньшую территорию. К примеру, на острове может не быть крупных рек, а саванны или леса могут иметь меньшую протяженность. На континентальном острове должны жить все группы животных, характерные для материка, от которого он отделился. Отсутствие на острове каких-то групп или видов объясняется либо тем, что они приурочены к биотопам, не имеющимся на острове, либо их вымиранием вследствие недостатка условий для поддержания необходимой численности популяций. К факторам, вызывающим островное обеднение, относится, естественно, и антропический.
На океанических островах могли укорениться лишь виды, способные активным или пассивным путем пересекать водные преграды. А поскольку такой способностью обладают далеко не все животные, бедность и дефектность фауны островов можно предсказать заранее. На океанических островах обитают летающие животные — птицы, летучие мыши, многие насекомые. Из нелетающих форм раньше других здесь появляются грызуны, из рептилий — гекконы. И те и другие могут заноситься на «плотах», т. е. плавающих стволах деревьев, вырванных или сломанных тропическими ураганами и выброшенных в море.
Даже крупные древние континентальные острова имеют относительно бедную и дефектную фауну. Так, на Мадагаскаре обитают только 2 семейства амфибий, но нет строго пресноводных рыб, сухопутных черепах из семейств трехкоготные и пресноводные, из змей отсутствуют аспидовые и гадюки и такие характерные для Африки семейства птиц, как страусы, птицы-секретари, цесарки, журавли, птицы-носороги, дятлы, вьюрковые и др. Из млекопитающих на Мадагаскаре нет и типично африканских семейств — слонов, носорогов, обезьян, кошек, гиен, антилоп, трубкозубов и т. д. Особенно следует подчеркнуть отсутствие копытных, кроме речной свиньи, и обилие хорошо летающих рукокрылых, родственных как африканским, так и индо-малай-ским формам.
Следовательно, бедность и дефектность фауны — общий и характерный признак всех островов, проявляющийся в тем большей степени, чем остров меньше по площади и дальше находится от материка или другого острова.
Эндемизм — второй характерный признак островной фауны — является результатом изоляции острова от других участков суши. Степень эндемизма тем больше, чем древнее остров и чем он более удален от материка. Таким образом, изоляция является причиной не только бедности и дефектности фауны, но и ярко выраженного островного эндемизма. Так, на Мадагаскаре, отдаленном от Африки морским проливом шириной 400 км и ставшем островом еще в мезозое, живут эндемичные семейства млекопитающих (тенреки из насекомоядных, 3 семейства лемуров) и птиц (нелетающие пастушковые куропатки, филепитты, ванги, а в прошлом и эпиорнисы), многочисленные эндемичные роды и огромное количество видов всех классов животных. Фауна птиц Новой Каледонии содержит более 20 % эндемичных видов и 1 эндемичное семейство.
Геологически молодые, расположенные близко от материка острова (например, Британские, Сахалин) эндемиков высокого ранга не имеют, хотя эндемичные подвиды или даже виды здесь обитают. Следовательно, процент эндемизма и систематический ранг эндемиков могут служить показателем геологического возраста острова. К примеру, согласно зоогеографическим данным, и Британия, и Сахалин стали островами только в четвертичном периоде, что подтверждается геологическими исследованиями.
Богатство фауны острова, степень ее эндемизма могут зависеть, кроме того, от его площади, разнообразия рельефа и других природных условий, а также от климатической зоны, в которой он находится. Все это создает большее или меньшее разнообразие экологических ниш.
Третья особенность островных фаун заключается в том, что они нередко являются последними убежищами архаичных видов животных. Такие виды немногочисленны и встречаются на Мадагаскаре, в Новой Зеландии, на Сулавеси, а также на Антильских островах. В одной из оригинальнейших наземных фаун — новозеландской — вообще отсутствуют аборигенные млекопитающие, но имеется древнейший вид рептилий — гаттерия, или туатара, а также гладконогие лягушки лиопельмы (2—3 вида) — древние и архаичные представители отряда бесхвостых. В состав современной мадагаскарской фауны входят древние тенреки (30 видов), настоящие лемуры, индри и руконожки (из примитивных приматов), легуаны (из игуановых рептилий). Интересно общее распространение последних. Легуаны представлены многочисленными видами в фаунах Северной и Южной Америки, а в Восточном полушарии встречаются на островах Фиджи и Тонга. На островах Куба и Гаити до нашего времени дожили архаичные щелезубы (2 вида), представители насекомоядных млекопитающих. В ископаемом состоянии близкие роды обнаружены в Северной Америке.
Исследователей фауны островов часто поражают ее необычные представители: например, с одной стороны, гиганты — гигантский варан на острове Комодо, черепахи на острове Маврикий, истребленные человеком птицы эпиорникс на Мадагаскаре и дронты на Реюньоне (также достигали гигантских размеров), с другой стороны — карлики, такие как мелкая форма тигра на Зондских островах, мелкие барсуки и лисицы на острове Сардиния и т. д. (ископаемые слоны на острове Мальта достигали размеров кабана). Островная популяция лошадей, предки которых были завезены с материка, выродились в мелкую породу — пони. Причины как одного, так и другого явления до настоящего времени не выяснены.
Немногочисленные, а то и одиночные виды животных, попав на острова, оказываются родоначальниками процветающих и достаточно многочисленных эндемичных групп. Этот феномен адаптивной радиации в рамках небольшого количества эндемичных групп представляет собой четвертую особенность островных фаун. Адаптивная радиация обусловливается наличием на островах разнообразных незанятых экологических ниш. Поэтому некоторые виды, попав на острова, настолько изменяются, что иногда уже мало чем напоминают предковую форму. Классическим примером адаптивной радиации являются гавайские птицы-цветочницы (семейство Drepanididae). Их предок, американский вьюрок, залетев на Гавайские острова, не нашел здесь конкурентов и дал начало нескольким эволюционным ветвям, нынешние представители которых по внешнему виду и образу жизни напоминают зябликов, медососов, пищух, дубоносов и дятлов.
ПРИНЦИПЫ И МЕТОДЫ
Дата добавления: 2017-10-09; просмотров: 2509;