Общая характеристика
Тело птиц состоит из округлой головы, туловища, начинающегося очень подвижной шеей, передних конечностей, превратившихся в крылья, и хорошо развитых ног. Форма тела — обтекаемая, лишенная выступающих частей, компактная;
крылья у птиц, передвигающихся по земле или находящихся в покое, сложены на спине. Поверхность распростертых во время полета крыльев значительно превышает поверхность туловища. Отмеченные особенности имеют большое значение для обеспечения передвижения птиц в воздухе, а также по земле. Хвост отсутствует.
Покровы. Кожа тонкая, что имеет существенное значение для уменьшения массы тела птиц и обеспечения подвижности перьев. В ней нет, как и у современных рептилий, желез, клетки наружных слоев ее эпидермиса сильно ороговевают, вследствие чего резко уменьшается испарение воды, столь необходимой для интенсивного обмена веществ, присущего птицам. Соедини-тельнотканный слой кожи подстилается рыхлой клетчаткой, в которой могут накапливаться запасы жира.
Тело покрыто перьями. Различают перья контурные и пуховые (рис. 99).
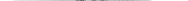
Рис. 99. Контурное перо (общий вид снизу):
1 — ствол (стержень); 2 — очин (на рисунке частично вскрыт); 3 — отверстие очина; 4 — душка (омертвевший сосочек); 5 — наружное опахало; 6 — внутреннее опахало; 7 — пуховая часть опахала; 8 — добавочный стержень
Контурное перо состоит из рогового стержня, который разделяется на очин и ствол, от ствола отходят длинные роговые пластинки — бородки первого порядка, от них — более короткие бородки второго порядка, сцепленные крючочками, в результате чего образуется прочная пластинка — опахало. В пуховом пере стержни слабые, бородки — мягкие, не сцепленные крючочками. Различают еще пух, в котором стержня нет, а имеются только мягкие бородки. Контурные перья прикреплены к коже не на всей поверхности, а только на определенных участках — птерилиях. Остальные участки кожи — аптерии — лишены контурных перьев, хотя они прикрыты ими. На этих участках у большинства пиц есть пух и пуховые перья. Последние могут быть и на птерилиях. Контурные перья названы так потому, что они в значительной степени определяют форму (контур) птиц. Они образуют прочную поверхность крыльев и на заднем конце тела — рулевую лопасть. Следовательно, контурные перья играют первостепенную роль в обеспечении полета. Весь перьевой покров предохраняет тело от излишней отдачи тепла и от механических повреждений, столь возможных во время полета, а также при передвижении по земле, на деревьях и т.д. Отмеченные положительные свойства перьевого покрова сочетаются с его малой массой, что так важно для летающих животных. Для сохранения эластичности контурных перьев птицы при помощи клюва смазывают их выделениями единственной кожной железы — копчиковой, расположенной около клоаки.
Образование пера начинается с выпячивания эктодермального сосочка, схожего с первыми стадиями развития роговых чешуи рептилий. Внутри сосочка имеется вырост соединительнотканно-го слоя кожи с кровеносными сосудами, питающими развивающееся перо. Снаружи сосочек окружен эпидермальным чехликом, под покровом которого происходит ороговение групп эктодер-мальных клеток и превращение их в бородки. По окончании развития бородок чехлик сбрасывается, освобождая готовое перо в виде пуха. При формировании пуховых и контурных перьев образуются стержни, от которых отходят бородки. Следовательно, готовые перья представляют собой чисто эктодермальные образования, в развитии которых принимает участие мезодермальный слой кожи.
Несмотря на достаточную прочность перьев, они все-таки повреждаются в результате воздействия разных внешних факторов и изнашиваются. Поэтому у птиц происходит периодическая смена оперения — линька. У одних птиц линьке одновременно подвергается весь перьевой покров. Такие птицы во время линьки не могут летать и прячутся в укромных местах. У других же птиц смена оперения совершается постепенно. Линька первого типа наблюдается у видов, которые добывают пищу не в полете; линька второго типа характерна для видов, которые настигают свою жертву на лету. Смена перьев необходима также в связи с изменениями условий среды в разные времена года, например, в зимнем оперении больше пуха и пуховых перьев, чем в летнем, и т. д.
Окраска перьев птиц разнообразна и характерна для каждого вида. Она обусловлена сочетанием разных пигментов: темных — меланинов, желтых и красных — липохромов. При отсутствии пигментов окраска перьев белая. Значение окраски различно: благодаря ей птицы менее заметны на фоне тех биотопов, где они обитают; она облегчает общение между особями одного и того же вида и в том числе встречу самцов и самок. Она меняется в зависимости от изменений абиотических условий среды, например, птицы, обитающие в более влажных и теплых местах их
ареала, имеют более темную окраску, а в более сухих и холодных местах — более светлую. У ряда видов окраска может меняться в связи с изменением сезонных климатических условий.
Нервная система. В связи с очень энергичным образом жизни птиц, потребовавшим усложнения всех систем органов, нервная система развита значительно лучше, чем у рептилий. Головной мозг намного превосходит по размерам мозг рептилий (рис. 100).

Рис. 100. Головной мозг голубя сбоку:
1 — большие полушария; 2 — обонятельные доли; 3 — эпифиз; 4 — гипофиз; 5 — хиазма и зрительные нервы; 6— зрительные доли среднего мозга; 7 — мозжечок;
8 — продолговатый мозг
Передний мозг велик, полосатые тела его, в которых находятся центры регуляции зрения, движения и других функций, весьма развиты. Кора полушарий выражена значительно лучше, чем у пресмыкающихся. В результате этих и других прогрессивных изменений роль переднего мозга как координатора всей сложной жизнедеятельности птиц резко возросла, способность к установлению новых связей со средой путем более быстрого приобретения условных рефлексов значительно улучшилась. Обонятельные доли развиты меньше, чем у ранее рассмотренных классов позвоночных, так как у птиц в их отношениях со средой главную роль играют зрение и слух. Усложнились и другие отделы головного мозга, в особенности мозжечок, имеющий очень крупные размеры. Значение его чрезвычай-f но возросло в связи с приспособлением птиц к полету. Спинной ; мозг ввиду редукции хвоста короткий, но строение его сильно , усложнилось. Головных нервов — 12 пар.
Органы чувств. Глаза велики, их масса у многих видов превосходит массу головного мозга. Дальность зрения у многих видов очень велика, поле зрения весьма широко; аккомодация к видению на разных расстояниях совершенна и осуществляется разными способами — изменением кривизны хрусталика, а также роговицы, в меньшей степени более старым способом, т. е. перемещением хрусталика. Птицы хорошо различают цвета, их оттенки, особенности рисунка и окраски окружающих их предметов. Необходимость столь совершенного зрения птиц вполне понятна, если учесть быстроту их передвижения во время полета, да и во многих случаях по земле.
Орган слуха устроен более сложно, чем у рептилий; нижний мешочек, воспринимающий звуковые колебания, увеличен за счет отходящего от него отростка (его называют завитком улитки, сравнивая с гомологичным органом млекопитающих, у которых этот отросток имеет 2,5 оборота); слуховая косточка благодаря изменениям в ее строении лучше передает в лабиринт звуковые колебания барабанной перепонки, расположенной в более развитом, чем у рептилий, наружном слуховом проходе; обрамление отверстия последнего перьями (особенно у совиных) напоминает слуховую раковину млекопитающих, играющую роль рупора. Птицы, как известно, издают разнообразные звуки, объединяющиеся у многих видов в сложные мелодии. В этом отношении птицы превосходят млекопитающих, способных издавать преимущественно однообразные звуки. Звуковая деятельность играет в жизни птиц огромную роль: в общении родителей с птенцами, самцов с самками, с другими особями того же вида, а также с представителями других видов.
Обоняние, как отмечалось, развито слабее, чем у других позвоночных, но у видов, питающихся падалью, которую они находят по запаху, оно развито лучше. Вкусовые раздражения воспринимаются особыми скоплениями клеток, расположенными на слизистой оболочке ротовой полости и языке. Но эти органы играют в жизни птиц несравненно меньшую роль, чем у млекопитающих. В коже имеются свободные окончания нервов и скопления разных чувствующих клеток, функции которых различны: осязание, восприятие изменений температуры, положения перьев и др.
Для нервной деятельности птиц характерны разнообразные и очень сложные инстинкты. Инстинктивная деятельность проявляется во взаимоотношениях самцов и самок, постройке гнезд, выкармливании и охране потомства, в способности многих птиц к дальним перелетам в связи со сменой времен года и т. д. Инстинкты проявляются на определенном этапе индивидуального развития птиц в определенных условиях внешней среды. Так, например, инстинкт ухаживания самцов за самками проявляется при достижении ими половой зрелости, наступление которой, в свою очередь, сильно зависит у ряда видов от удлинения светлого периода суток весной. Другие инстинкты свойственны птицам в течение всей жизни. Отсутствие сознания в инстинктивных действиях можно проиллюстрировать на следующем примере. Хорошо известно, что кукушки подбрасывают свои яйца в гнезда мелких воробьиных птиц. Вылупившийся кукушонок, еще слепой и глухой, выбрасывает яйца и птенцов хозяев гнезда, а те продолжают кормить быстро растущего, прожорливого нахлебника. Способность к образованию условных рефлексов у птиц развита во много раз лучше, чем у пресмыкающихся. Этой способностью люди пользуются при разведении домашних птиц. У кур, например, легко и быстро вырабатываются условные рефлексы: они сбегаются к месту кормления, вечером возвращаются на насесты и т. д.
Инстинктивная деятельность особенно ярко проявляется у птиц, совершающих дальние сезонные перелеты: весной — из южных стран в северные страны, где они размножаются и выкармливают своих птенцов, а осенью с приближением более холодного времени — обратно в южные страны. Инстинктивный характер перелетов подтверждается тем, что молодые птицы, появившиеся на севере, в ряде случаев летят на юг самостоятельно, а не с родителями, т. е. не используя опыта последних. Были поставлены опыты с перелетными птицами в условиях, которые оставались характерными для летнего времени года, тем не менее подопытные животные с приближением осени испытывали беспокойство и пытались улететь. Пути перелетов, места пребывания перелетных птиц в южных странах и места, где они выводят свое потомство в северных странах, почти всегда постоянны. Проблема перелетов до сих пор остается до конца не разрешенной, в частности, не выяснено с достаточной полнотой, чем руководствуются птицы, совершая столь далекие передвижения, хотя доказано, что они могут учитывать положение солнца, луны и звезд, обладают «компасным чувством» и т. д. Взрослые птицы, возвращающиеся в южные страны (причем в течение ряда лет), используют свой опыт в выборе наиболее удобных мест для миграций (вдоль рек, границ лесов и т.д.). Во время перелетов большое число птиц, конечно, погибают. Однако поскольку перелеты продолжаются в течение огромных промежутков времени, это означает, что они имеют большое приспособительное значение, так как обеспечивают размножение птиц в благоприятных условиях. Таким образом, способность к таким миграциям возникла у птиц в результате естественного отбора в течение длительного времени. Формирование перелетов начиналось с кочевок на небольшие расстояния. Постепенно расстояние увеличилось, когда это оказалось выгодным для сохранения вида. В заключение следует подчеркнуть, что известен ряд фактов, подтверждающих некоторую изменчивость миграций птиц, что является основой для возникновения новых путей перелетов. Понятно, что способность птиц к таким длительным миграциям стала возможной благодаря их совершенному приспособлению к полету и прогрессивным особенностям всех систем органов, в первую очередь нервной системы. Птицам свойственны и некоторые проявления рассудочной деятельности. Так, например, некоторые хищные птицы поджидают свою жертву, скрывшуюся в кусты, с противоположной стороны убежища.
Скелет. Череп в связи с большим размером головного мозга относительно велик. Он соединен с позвоночным столбом, как у тех пресмыкающихся, от которых произошли птицы, одним мыщелком — затылочным. Кости черепа тонкие, что уменьшает массу головы и облегчает ее движения при захвате добычи и во время полета, а также понижает центр тяжести тела, способствуя большей его устойчивости. Защита мозга усиливается сращением у взрослых птиц костей черепа, хотя у зародышей швы, разделяющие кости, имеются. В состав черепа входят в основном те же кости, что и у пресмыкающихся. Квадратные кости подвешивают, как и у земноводных и пресмыкающихся, нижнюю челюсть (образованную несколькими костями) к мозговому отделу черепа. Нёбо птиц образовано отростками верхнечелюстных костей, нёбными костями, сошником и крыловидными костями (рис. 101).
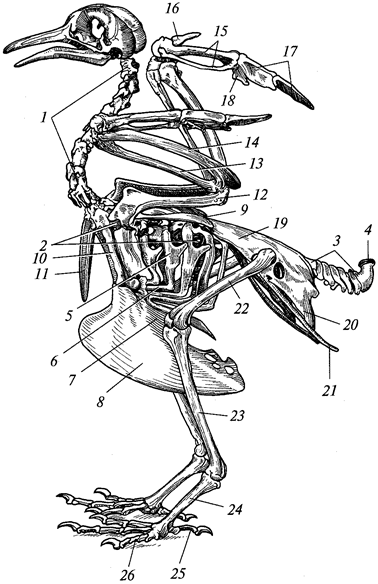
Рис. 101. Скелет голубя:
1 — шейные позвонки; 2 — грудные позвонки; 3 — хвостовые позвонки; 4 — копчиковая кость; 5 — спинная часть ребра с крючковидным отростком; 6 — брюшная часть ребра; 7 — грудина; 8 — киль грудины; 9 — лопатка; 10 — коракоид; 11 — ключица; 12 — плечо; 13 — лучевая кость; 14 — локтевая кость; 15 — пясть; 16 — I палец; 77—11 палец; 18 — III палец; 19 — подвздошная кость; 20 — седалищная кость; 21 — лобковая кость; 22 — бедро; 23 — голень; 24 — цевка; 25 — I палец; 26 — IV палец
Для всех отделов позвоночника, кроме шейного, характерно сращение многих позвонков между собой. Эта особенность позвоночного столба обусловливает неподвижность туловища птиц, что имеет большое значение при полете. Шейный отдел позвоночника очень длинный. Строение первых двух шейных позвонков (атланта и эпистрофея) и седлообразные суставные поверхности остальных шейных позвонков обеспечивают чрезвычайную гибкость шеи. Поэтому голова птиц очень подвижна, что облегчает поиски и хватание пищи, чистку перьев, смазывание их выделениями копчиковой железы и т. д.
Ребра состоят из двух частей — позвоночной и грудинной. Такое строение ребер облегчает изменение объема грудной клетки, что очень важно при вдохе и выдохе. Соседние ребра соединены между собой маленькими клювовидными отростками, благодаря чему усиливается прочность грудной клетки. Грудина у летающих птиц и у пингвинов, плавающих при помощи ластов, т. е. видоизмененных крыльев, имеет киль, к которому прикрепляются сильно развитые большие грудные и другие мышцы. У бегающих птиц (страусов и др.) киля нет.
Передние конечности у птиц превратились в крылья. Основу крыла составляет измененный скелет передних конечностей наземных рептилий, который состоит из хорошо развитой плечевой кости, прямой и более тонкой лучевой кости, изогнутой и более толстой локтевой кости и кисти, в которой произошли редукция ряда костей и слияние некоторых костей между собой. Из костей запястья остались лишь две косточки. Пястный отдел кисти представляет собой две кости, соединенные своими концами, образующие «пряжку». Сохранились рудименты трех первых пальцев, из которых второй развит лучше других. В результате этих изменений передняя конечность потеряла способность к передвижению по земле и не может выполнять хватательные функции, но ее кости образуют удлиненное и надежное основание для прикрепления мышц и перьев. Плечевой пояс состоит из узких лопаток, вороньих костей и ключиц. Последние срастаются нижними концами и образуют «вилочку».
Вследствие превращения передних конечностей в крылья движение по земле осуществляется только при помощи задних конечностей, причем вся тяжесть падает на них. В связи с этим скелет таза испытал ряд существенных изменений. Таз птиц очень велик. У рептилий он был соединен с позвоночником в области двух (крестцовых) позвонков. У птиц сращение таза с позвоночным столбом, кроме крестцовых позвонков (их тоже обычно два), распространяется на оставшиеся хвостовые, все поясничные и даже задние грудные позвонки. Эти позвонки срастаются между собой и образуют сложный крестец, в состав которого может входить до 24 позвонков. Таким образом, ноги птиц имеют очень прочную опору. Седалищные кости срастаются на всем протяжении с подвздошными. Лобкового соединения нет и таз спереди широко открыт, благодаря чему птицы могут нести яйца большой величины.
С тазом сочленяется бедро. После бедра идет очень длинная голень, состоящая из большой берцовой кости, к которой в виде тонкого рудимента прирастает малая берцовая кость. Как показали эмбриологические исследования, с дистальным концом большой берцовой кости срастаются слившиеся кости первого (про-ксимального) ряда — предплюсны. Второй (дистальный) ряд костей предплюсны срастается с плюсневыми костями и образуется довольно длинная кость — цевка. В результате описанных изменений скелет ноги птицы стал более прочным, а общая длина этой конечности значительно удлинилась, что обеспечило более быстрое передвижение и облегчило добывание пищи при помощи клюва. Пальцев у большинства птиц четыре, причем первый направлен назад.
Скелет птиц в связи с приспособлением их к полету довольно сильно облегчен, что достигается разными способами: многие кости очень тонкие, в них мало костного мозга, в большие кости заходят ответвления воздушных мешков и т. д. Кости, наполненные воздухом, называются пневматическими.
Мышечная система. Она значительно дифференцированна и массивнее, чем у рептилий. Мышцы в основном сосредоточены на туловище и проксимальных частях конечностей, что способствует усилению компактности тела, важной для полета. Мышцы, выполняющие наибольшую нагрузку, содержат белок миоглобин (мышечный гемоглобин, близкий по составу к гемоглобину эритроцитов), благодаря которому в их волокнах накапливается резерв кислорода. Многочисленные мышцы шеи обеспечивают разнообразные и быстрые движения головы. Наибольшего развития достигает большая грудная мышца, прикрепляющаяся к килю грудины.
В функции этой мышцы входит опускание крыла, для чего необходимо преодолевать большое сопротивление воздуха. Масса ее может доходить до 1/5—1/4 массы тела. Подъем крыла обеспечивается малой грудной (подключичной) мышцей, которая прикреплена к вороньей кости (коракоиду) и грудине, а ее сухожилие — к головке плечевой кости. Расширение грудной клетки происходит благодаря сокращению межреберных и некоторых других мышц. Очень хорошо развиты мускулы ног, из которых самые большие прикреплены начальными концами к разным участкам таза, а более мелкие находятся на бедре и голени. Весьма совершенны разгибатели и сгибатели пальцев. У птиц развился мышечный механизм, обеспечивающий им прикрепление при помощи пальцев к веткам деревьев во время сна. Имеется подкожная мускулатура, от которой зависит изменение положения перьев на больших участках поверхности тела и отдельных перьев.
Различают два главных вида полета — парящий и машущий. Первый присущ многим птицам: обладая прекрасно развитой нервной системой, они хорошо ориентируются в воздушных течениях и используют восходящие токи воздуха для парения при помощи распростертых крыльев, редко взмахивая ими. Второй возможен только при непрерывной работе крыльев, число взмахов которых у самых мелких птиц — колибри доходит до 50—80 в секунду. Скорость полета различна: у голубей — 30—60 км/ч, у ласточек — 40— 60, у стрижей — до 100—120 км/ч. Скорость полета возрастает при попутном ветре и замедляется при встречном. Перелетные птицы, хорошо приспособленные к длительному нахождению в воздухе, преодолевают в сутки в среднем 200—400 км. Полет происходит, как правило, на небольшой высоте (100—300 м), но птицы могут подниматься и на несколько тысяч метров. Длина пролетных путей некоторых видов птиц достигает 12000 км.
Пищеварительная система. Все современные птицы не имеют зубов. Края челюстей — роговые, образуют клюв, служащий для захвата пищи и у некоторых видов — для размельчения ее. Поскольку в ротовой полости птиц не происходит механической обработки пищи, как это имеет место у млекопитающих, эта обработка почти целиком совершается в других частях пищеварительной трубки. В ротовую полость открываются протоки слюнных желез, секрет которых смачивает пищу. Из ротовой полости пища попадает в глотку и далее в пищевод, где у многих птиц она задерживается в особом расширении — зобе. В нем смоченная пища размягчается. Из зоба через нижний отдел пищевода пища проходит в железистый желудок, где она обволакивается пищеварительными соками, а затем переходит в мышечный желудок. У хищных птиц этот отдел выражен слабо, но он прекрасно развит у растительноядных видов. Мощная мускулатура желудка, его жесткая внутренняя поверхность, а также находящиеся в его полости проглоченные камешки обеспечивают перемалывание различных пищевых продуктов, заключенных в крепкие оболочки. Здесь же, поскольку пища в предыдущем отделе была смешана с пищеварительными соками, продолжается ее химическое разложение, но в основном оно происходит в тонком отделе кишечника.
Тонкая кишка развита лучше, чем у пресмыкающихся, и в 2— 4 раза превосходит длину тела, но она значительно короче, чем у многих млекопитающих. Очень длинная тонкая кишка и содержащаяся в ней пища чрезмерно увеличивали бы массу тела. Сравнительно небольшая длина тонкого кишечника возмещается сильным действием пищеварительных соков. Эти соки выделяются железами кишечной стенки и хорошо развитой поджелудочной железы. Важную роль в пищеварении играет печень, состоящая у птиц из двух долей. Желчного пузыря у некоторых птиц (например, у голубя) нет. Всасывание переваренной пищи происходит очень быстро благодаря наличию на внутренней поверхности тонкой кишки большого количества ворсинок.
Толстая кишка очень короткая, она заканчивается клоакой, в которую впадают, как и у пресмыкающихся, выводные протоки мочевых и половых органов. В короткой толстой кишке не могут задерживаться и формироваться экскременты, и птицы испражняются очень часто. Быстрое опорожнение кишечника, возможное благодаря интенсивным процессам пищеварения у птиц, выгодно для облегчения их массы.
Состав пищи различен не только у разных видов, но и у птенцов и взрослых особей одного и того же вида. Так, например, куриные зерноядны, но их птенцы питаются насекомыми. Очевидно, для роста и развития молодых птиц большое значение имеет животный корм.
Дыхательная система. Она устроена значительно сложнее, чем у рептилий. Бронх, ответвляющийся от заднего конца трахеи, входит в легкое, пронизывает его насквозь, и его полость переходит за пределами легкого в полость большого воздушного мешка. От центрального бронха отходят вторичные бронхи, из которых несколько бронхов (чаще четыре) тоже пронизывают легкое и соединяются с другими воздушными мешками. Остальные вторичные бронхи (10—14) заканчиваются слепо, т.е. не соединены с воздушным мешком. Вторичные бронхи соединены между собой многочисленными парабронхами, от которых отходит множество каналов — бронхиол, густо оплетенных капиллярами. Поверхность бронхиол очень велика и обеспечивает интенсивный газообмен, столь необходимый птицам.
Воздушные мешки — тонкостенные образования — отходят от упомянутых выше сквозных бронхов (рис. 102). Имеются следующие воздушные мешки: парные — шейные, подмышечные, переднегрудные, заднегрудные и брюшные, непарный — межключичный.
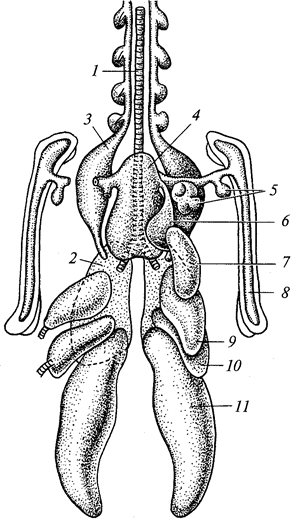
Рис. 102. Схема воздушных мешков птиц; вид с брюшной стороны:
1 — трахея; 2 — легкое; 3 — шейный мешок; 4 — межключичный мешок; 5—8 — выросты межключичного мешка; 9 — переднегрудной мешок; 10 — заднегрудной мешок; 11 — брюшной воздушный мешок
У птицы, не находящейся в полете, при вдохе вследствие сокращения определенных мышц расширяется грудная клетка (при этом поднимается грудная кость), что влечет за собой расширение воздушных мешков и приток в них через легкие воздуха. Сами легкие малоэластичны и почти не расширяются. При выдохе, происходящем при сжатии грудной клетки и опускании грудины, воздушные мешки сжимаются и значительная часть наполняющего их воздуха попадает в легкие. Так как в воздушных мешках кислород не потребляется, то капилляры бронхиол насыщаются кислородом не только при вдохе, но и при выдохе. Поэтому принято говорить, что у птиц двойное дыхание. У летящих птиц грудина остается неподвижной (ибо она является опорой для главных летательных мышц), расширения и сжатия грудной клетки не происходит и вентиляция легких в таком состоянии совершается в результате того, что при подъеме крыльев воздушные мешки расширяются, а при опускании крыльев сжимаются.
Интенсивность газообмена усиливается у птиц благодаря очень высокой частоте вдохов и выдохов (до 90-100 раз в минуту у небольших птиц, т.е. значительно больше, чем у млекопитающих).
Кроме того, гемоглобин птиц быстрее отдает тканям кислород, чем гемоглобин млекопитающих.
Воздушные мешки помимо их важнейшей роли в процессах дыхания выполняют и другие функции: заполняя брюшную полость тела и проникая в некоторые длинные кости, они уменьшают приблизительно на 10 % массу тела, являются изолирующей прокладкой между органами, уменьшают теплоотдачу, облегчают процессы дефекации и откладки яиц.
У птиц очень длинная трахея. Просвет ее поддерживается хрящевыми кольцами, которые в некоторых случаях окостеневают. Гортань, от которой начинается трахея, у птиц не приспособлена к издаванию звуков, а певчей, или нижней, гортанью является орган, возникший в результате преобразования нижней части трахеи и начальных частей главных бронхов, имеющий внутри перепонки и сложно-дифференцированную мускулатуру, способствующую вибрации последних. Трахея помимо очищения воздуха от пыли и согревания его имеет значение для усиления издаваемых нижней гортанью разнообразных звуков.
Кровеносная система.Сердце птиц четырехкамерное и состоит из двух предсердий и двух желудочков (рис. 103). Правый желудочек, получающий венозную кровь из правого предсердия, полностью отделен от левого желудочка, получающего артериальную кровь из левого предсердия. Левая дуга аорты атрофировалась, а правая дуга аорты, отходящая от левого желудочка, несет артериальную кровь, попадающую по разветвлениям аорты, во все части тела. Следовательно, у птиц полностью отделился артериальный ток крови от венозного, и их организм снабжается только артериальной кровью. Это способствовало резкому усилению обмена веществ у птиц по сравнению с их предками.
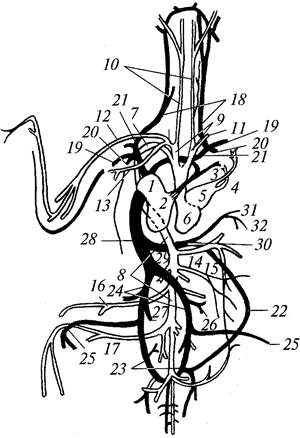
Рис. 103. Кровеносная система птиц (светлое — сосуды с
артериальной кровью; черное — с венозной):
1 — правое предсердие; 2 — правый желудочек; 3 — легочная артерия; 4 — легочные вены; 5 — левое предсердие; 6 — левый желудочек; 7 — правая дуга аорты; 8 — спинная аорта; 9 — безымянная артерия; 10 — сонная артерия; 11 — подключичная артерия; 12 — плечевая артерия; 13 — грудная артерия; 14 — внутренностная артерия; 15 — брыжеечная артерия; 16 — бедренная артерия; 17 — седалищная артерия; 18 — яремная вена; 19 — плечевая вена; 20 — грудная вена; 21 — передняя полая вена; 22 — копчиково-брыжеечная вена; 23 — воротная вена почки; 24 — общая подвздошная вена; 25 — седалищная вена; 26 — бедренная вена; 27— почечная вена; 28 — задняя полая вена; 29 — печеночная вена; 30 — воротная вена печени; 31 — поджелудочная вена; 32 — брыжеечная вена
Сердце птиц очень часто сокращается: у мелких видов в состоянии покоя — 400—600 раз в минуту, в полете — до 1000, у видов средней величины в состоянии покоя — 200-300, в полете — до 600 раз в минуту; у более крупных видов частота сокращений меньше, но она превосходит аналогичные показатели рептилий и даже млекопитающих. Следовательно, кровь движется в теле птиц с очень большой скоростью, что способствует быстрому удалению продуктов диссимиляции и насыщению тела кислородом и питательными веществами. Сердце птиц, выполняющее такую большую работу, относительно велико: масса его достигает у мелких видов 2,4—2,85 % всей массы тела; у более крупных видов относительная масса сердца меньше, но она значительно больше, чем у рептилий (0,2—0,3 %) и даже у млекопитающих (0,2—1,0 %). Количество крови, число эритроцитов и содержание гемоглобина у птиц значительно больше, чем у рептилий. Отмеченные особенности кровеносной системы, а также ряд других, не упомянутых здесь, обеспечивают весьма энергичный образ жизни птиц.
Выделительная система. В результате интенсивного обмена веществ у птиц выделяется большое количество продуктов диссимиляции, которые как можно скорее должны быть удалены из организма. Эту функцию выполняют тазовые почки. Последние, унаследованные птицами от рептилий, усложнились, относительная масса их сильно увеличилась, достигнув 1-2,6 % массы всего тела. Почки у большинства видов птиц внешне подразделяются на три доли. Моча удаляется через мочеточники в клоаку, она, как и у пресмыкающихся, богата мочевой кислотой и консистенция ее кашицеобразная. Мочевого пузыря у птиц нет, и моча часто удаляется из клоаки. Накопление ее в мочевыносящих путях увеличивало бы массу тела птиц.
Размножение. Мужские половые органы сходны с теми же органами пресмыкающихся. Семенники лежат около почек. Левый семенник обычно больше развит, чем правый, но семя вырабатывается обеими железами. Семяпроводы, образовавшиеся, как и у пресмыкающихся, из вольфовых каналов, открываются в клоаку.
Оплодотворение внутреннее, хотя специальных совокупительных органов у большинства птиц нет и передача семени в половые пути происходит при половом акте в результате прижимания клоаки самца к клоаке самки.
Женская половая система почти у всех птиц состоит только из одного левого яичника и левого яйцевода (бывшего мюллерова канала). Правый яичник развивается в очень редких случаях, а производимые им яйцеклетки выходят через левый яйцевод.
Редукция на одной стороне тела женских половых органов произошла, вероятно, в связи с необходимостью облегчения массы тела, меньшее же количество откладываемых птицами по сравнению с пресмыкающимися яиц компенсируется хорошо развитой заботой о потомстве. Созревшие яйцеклетки, окруженные желтком, попадают в воронку яйцевода. Яйцевод состоит из нескольких отделов: трубки, внутренняя поверхность которой покрыта мерцательным эпителием и имеет железы невыясненного значения; широкой части, стенки которой выделяют белок, и хлгазы (спиральные завитки на обоих полюсах яйца); узкой части, в которой выделяется тонкая белковая оболочка яйца; хорошо развитой, очень мускулистой матки, где яйцо окружается скорлупой; мускулистого влагалища, выталкивающего готовое яйцо в клоаку. Яйцеклетки оплодотворяются в верхней части яйцевода.
Яйца птиц разнообразны по размерам, окраске и толщине скорлупы. Обычно величина яиц пропорциональна размерам тела птиц, но отступления от этого правила нередки. Яйца, откладываемые на землю, камни и т. д., имеют более толстую скорлупу, чем яйца, откладываемые в гнезде. Яйца многих птиц имеют защитную окраску. Количество яиц в кладке у разных видов варьирует.
Половой диморфизм у ряда видов проявляется слабо (совы, голуби и др.), но у большинства птиц самцы отличаются от самок яркостью оперения, размерами, голосом, наличием хохла, воротника, различных роговых выростов. Самки же этих видов имеют неяркую окраску и лишены различных придатков. Способность к пению тоже присуща, как правило, самцам. В период спаривания половое возбуждение самцов может проявляться кроме пения различного рода звуками, телодвижениями и т. п.
Развитие оплодотворенных яиц начинается еще в половом аппарате самки, однако вскоре оно приостанавливается и продолжается уже после откладки яиц. Почти все птицы насиживают яйца, т. е. обогревают их своим телом. Обычно насиживают самки, но у ряда видов эту функцию выполняют оба родителя или только самцы. Продолжительность эмбрионального развития у разных птиц различна и характерна для каждого вида: у кур — 21 день, у самых больших из современных птиц, африканских страусов, — 55—60 дней, а у самых маленьких, колибри, — 10—12 дней.
Птицы относятся к амниотам, и их зародыши имеют те же оболочки, что и пресмыкающиеся, т. е. амнион, серозу и аллан-тоис. Когда у эмбриона развиваются легкие, аллантоис становится ненужным и ссыхается. Развившийся птенец пробивает скорлупу при помощи особого выроста на клюве — яйцевого зуба.
У одних птиц вылупившиеся птенцы вполне развиты и почти сразу начинают самостоятельно двигаться и добывать себе корм. Такие птицы называются выводковыми. К ним относятся, как правило, птицы, устраивающие свои гнезда на земле, у воды и т. д. (например, куриные, гусиные и др.). У других птенцы вылупляются недоразвитыми: слепые, голые или покрытые редким пухом, беспомощные — некоторое время их выкармливают родители. Такие птицы называются птенцовыми. К ним относятся, как правило, птицы, устраивающие свои гнезда на деревьях, вообще высоко над землей (например, голубиные, дятлы, воробьиные и др.). Между обеими группами нет резкой границы.
Происхождение. Птицы, как отмечалось выше, произошли от пресмыкающихся. Наиболее убедительным доказательством этой научной гипотезы в течение долгого времени считался отпечаток животного (обнаруженный в юрских отложениях), названного археоптериксом (Archaeopterix lithographica). В организации этой древней птицы сочетались признаки рептилий (длинный хвост, пальцы с когтями на передней конечности, наличие самостоятельной малой берцовой кости, зубы на челюстях и др.) и птиц (перьевой покров, примитивные крылья, цевка, большие глазницы и др.). Археоптериксы были величиной с сороку, они вели древесный образ жизни, передвигались, цепляясь когтями за ветки и, вероятно, планируя при помощи крыльев, усаженных большими перьями, могли перебираться с одного дерева на другое.
В последнее время появились сведения о нахождении остатков еще более древнего предка птиц, у которого признаки рептилий (в особенности сильные задние ноги) были выражены сильнее, чем у археоптерикса. Этим подтверждается гипотеза о происхождении птиц от небольших пресмыкающихся — динозавров, которые передвигались при помощи прыжков, опираясь на задние ноги. В меловом периоде мезозойской эры появились птицы, вполне похожие на современных, но у которых еще сохранились зубы. В начале третичного периода кайнозойской эры зубатых птиц уже не было. Таким образом, класс птиц подразделяется на два подкласса: ящерохвостые, или древние птицы (Archae-ornithes), к которым относятся археоптерикс и веерохвостые, или настоящие птицы (Neornithes) (к последним принадлежат и зубатые птицы, хотя некоторые орнитологи выделяли их в отдельный подкласс).
Дата добавления: 2017-10-09; просмотров: 1375;