Молочнокислое брожение
Молочнокислое брожение глюкозы является основным процессом при изготовлении заквасок, сыра и кисломолочных продуктов, а молочнокислые бактерии — наиболее важной группой микроорганизмов для молочной промышленности. Процесс молочнокислого брожения каждой гексозы молочного сахара выражается суммарным уравнением
С6Н12О6 + 2ФН + 2АДФ ¾¾¾¾® 2С3Н6О3 + 2АТФ + Н2О.
Наряду с молочной кислотой могут образовываться и побочные продукты брожения.
Молочнокислые бактерии по характеру продуктов сбраживания глюкозы относят к гомоферментативным или гетероферментативным. Гомоферментативные бактерии, как показывает их название, образуют главным образом молочную кислоту (более 90%) и лишь незначительное количество побочных продуктов. Гетероферментативные бактерии около 50% глюкозы превращают в молочную кислоту, а остальное количество — в этанол, уксусную кислоту и СО2. Однако провести резкую границу между гомо- и гетероферментативными молочнокислыми бактериями по образующимся продуктам брожения иногда бывает трудно. Так, отмечены факты образования отдельными штаммами гомоферментативных молочнокислых бактерий от 8 до 30% побочных продуктов, а гетероферментативные бактерии под воздействием ряда факторов могут вести себя как гомоферментативные.
Более характерным признаком при делении молочнокислых бактерий на группы является путь сбраживания глюкозы. Гомоферментативные бактерии сбраживают глюкозу по гликолитическому пути Эмбдена-Мейергофа-Парнаса, гетероферментативные — пентозофосфатным путем.
Гомоферментативное молочнокислое брожение. Механизм гомоферментативного молочнокислого брожения глюкозы к настоящему времени изучен подробно. Превращение глюкозы в пировиноградную кислоту идет в результате последовательных реакций гликолиза (см. стр. 101). В заключительной реакции пировиноградная кислота восстанавливается до молочной кислоты под действием фермента лактатдегидрогеназы. Роль восстановителя при этом играет НАДН.Н+, образовавшийся в реакции окисления 3-фосфоглицеринового альдегида (см. рис. 4.8).
Следовательно, при гомоферментативном молочнокислом брожении из 1 моля глюкозы образуются 2 моля молочной кислоты (с одновременным синтезом 2 моля АТФ).
При сбраживании глюкозы молочнокислыми бактериями могут образоваться два оптических изомера молочной кислоты: левовращающий D(-)-лактат и правовращающий L(+) лактат:
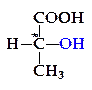 D(+)-лактат
D(+)-лактат 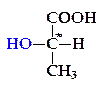 L(-)-лактат
L(-)-лактат
Изомеры отличаются расположением водородных атомов и гид-роксильных групп у асимметрического атома углерода, отмеченного звездочкой, — первый лактат имеет группу ОН справа и называется D-изомером (от лат. dexter — правый), второй имеет ее слева и поэтому назван L-изомером (от лат. laevus — левый).
L-лактат является промежуточным продуктом обмена веществ в человеческом организме и легко превращается в пировиноградную кислоту, которая окисляется до СО2 и Н2О в цикле трикарбоновых кислот или используется для синтеза гликогена. D-лактат, наоборот, очень медленно распадается в организме, а новорожденные его вовсе не утилизируют. Поэтому продукты для вскармливания грудных детей не должны содержать D-форму молочной кислоты или содержать ее мало. Следовательно, для приготовления детских кисломолочных смесей следует сочетать штаммы микроорганизмов, образующие L(+) или DL-молочную кислоту, например, бифидобактерии и ацидофильную палочку (а при производстве йогурта — термофильный стрептококк и болгарскую палочку).
Молочная кислота может быть не единственным конечным продуктом гомоферментативного молочнокислого брожения. Ход брожения часто изменяется в зависимости от конкретных условий (наличия СО2, кислорода, рН, температуры среды и т.д.). Возможные метаболические пути превращения глюкозы гомоферментативными молочнокислыми бактериями (а также другими бактериями и дрожжами) представлены на рисунке 5.1. Как видно из приведенной схемы, в качестве побочных продуктов могут образовываться летучие и нелетучие органические кислоты, глицерин, ацетальдегид, спирты, ацетон, ацетоин, диацетил, диоксид углерода и др.
Например, некоторые виды молочнокислых бактерий сбраживают часть глюкозы в этанол, уксусную и муравьиную кислоты (рисунок 5.1, путь б). Сначала глюкоза превращается в пировиноградную кислоту, которая затем расщепляется на ацетил-КоА и муравьиную кислоту (последняя может распадаться на СО2 и Н2). Далее половина молекул ацетил-КоА переходит в уксусную кислоту, другая половина — в этанол.
Данный путь сбраживания глюкозы характерен для таких нетипичных молочнокислых бактерий, как кишечная палочка Е. coli. Суммарная реакция брожения глюкозы этими бактериями имеет следующий вид:
C6H12O6+Н2O + 3ФН + 3АДФ ¾®
¾® С2Н5ОН + CH3COOH + 2Н2 + 2СО2 + 3АТФ
Часть глюкозы может превращаться в янтарную, пропионовую, яблочную и другие органические кислоты (путь в). Могут образовываться ацетон, изопропиловый и бутиловый спирты, а также масляная кислота (путь г).
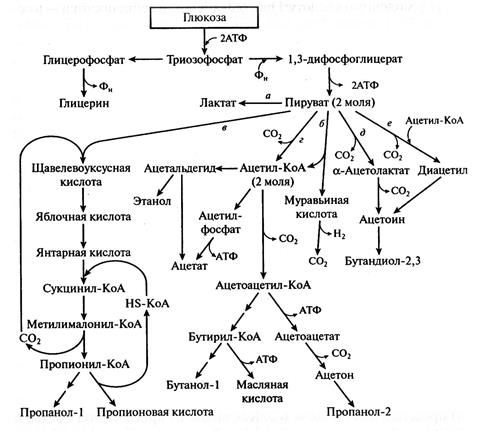
Рисунок 5.1 - Возможные варианты превращения глюкозы по пути Эмбдена-Мейергофа-Парнаса
(по Мецлеру, с изменениями)
Возможно также превращение глюкозы в ароматические вещества: ацетоин, бутандиол-2, 3-диацетил (пути д и е). Однако эти вещества образуются из глюкозы лишь в небольших количествах. Более значительное их количество образуется при сбраживании лимонной кислоты.
Гетероферментативное молочнокислое брожение. Бактерии группы лейконостоков и гетероферментативные молочнокислые палочки не могут сбраживать глюкозу по гликолитическому пути, для ее расщепления они используют пентозофосфатный (фосфоглюконатный, или гексозомонофосфатный шунт) путь. Это объясняется отсутствием у них ключевого фермента альдолазы, необходимого для расщепления фруктозо-1,6-дифосфата на две молекулы триозофосфата. Пентозофосфатный путь окисления глюкозы представлен на рисунке 5.2.

Рисунок 5.2 – Пентозофосфатный путь расщепления глюкозы
гетероферментативными молочнокислыми бактериями
Сначала происходит дегидрирование глюкозо-6-фосфата, катализируемое дегидрогеназой (глюкозо-6-фосфат—дегидрогеназой), которая в качестве акцептора водорода использует НАДФ+ или НАД+.
Затем 6-фосфоглюконовая кислота подвергается окислительному декарбоксилированию (при участии фермента 6-фосфоглюконат—дегидрогеназы) с образованием рибулозо-5-фосфата и восстановленной формы НАД или НАДФ. Рибулозо-5-фосфат под действием изомеразы далее превращается в рибозо-5-фосфат. Образовавшийся пен-тозофосфат при участии фосфокетолазы расщепляется на 3-фосфоглицериновый альдегид и ацетилфосфат, которые соответственно превращаются в лактат и этанол.
В ходе реакций по пентозофосфатному пути из каждого моля глюкозы образуются моль молочной кислоты, моль этанола и СО2:
C6H12O6 + ФН + АДФ ¾® С3Н6О3 + C2H5OH + СО2 + АТФ
Энергетический баланс брожения глюкозы, протекающего по этому пути, составляет одну молекулу АТФ. В аэробных условиях возможно образование двух молекул АТФ, тогда ацетил-фосфат превращается не в этанол, а в уксусную кислоту (см. рис. 5.2).
Фруктозо-6-фосфатный путь расщепления глюкозы бифидобактериями. Бифидобактерии в отличие от пентозофосфатного пути превращают глюкозу не в глюкозо-6-фосфат, а во фруктозо-6-фосфат, который далее расщепляется фосфокетолазой на эритрозо-4-фосфат и ацетилфосфат (рисунок 5.3). Затем эритрозо-4-фосфат и фруктозо-6-фосфат под действием ферментной системы перестройки Сахаров превращается в два моля пентозофосфата (ксилулозо-5-фосфата и рибозо-5-фосфата). Далее пентозофосфат расщепляется на ацетил-фосфат и глицеральдегид-3-фосфат, который переходит в пируват.
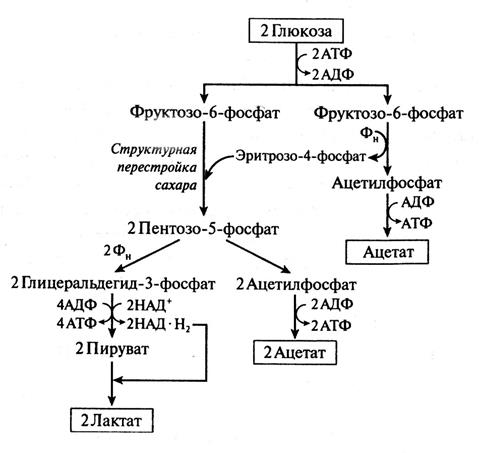
Рисунок 5.3 – Путь сбраживания глюкоза бифидобактериями
На последней стадии пируват восстанавливается до лактата (или расщепляется до ацетата и формиата). Таким образом, образование уксусной и молочной кислот происходит в молярном отношении 3:2. Выход АТФ составляет 21/2 моля на 1 моль глюкозы. Цитраты бифидобактерии не используют.
Бифидобактерии имеют низкую b-галактозидазную активность и плохо развиваются в молоке. Для стимуляции их роста целесообразно в закваски вводить культуры термофильного стрептококка обладающего повышенной b-галактозидазной активностью.
Дата добавления: 2017-08-01; просмотров: 3450;