Роль базальных ганглиев в индивидуально приобретенных реакциях.
Мы выше пришли к заключению, что условнорефлекторная деятельность исчезает после полной декортикации и разрушения базальных ганглиев и, наоборот, сохраняется после удаления одного неокортекса, при сохранности палеокортекса и базальных ганглиев. Спрашивается, какова роль базальных ганглиев в этих реакциях? Как уже указывалось (гл. VIII), базальные ганглии признаются многими авторами как передаточные станции от коры к промежуточному мозгу. Между тем, как показывают исследования Когана, базальные ганглии должны играть важную роль и в передаче возбуждения в центростремительном направлении. Именно Коган (7) при изучении электрической активности хвостатого тела отметил, что отводимые участки реагировали одинаковыми электрическими эффектами на различные раздражения даже противоположного характера, такие, например, как пищевые и болевые раздражения. Характерно, что эти эффекты возникали и при условных пищевых и оборонительных сигналах. Поэтому Коган находит, что эти подкорковые образования не содержат интегрирующих механизмов сложных поведенческих реакций. По его мнению, они являются передаточными станциями на центростремительных путях к интегрирующим механизмам этих реакций, которые находятся в гипоталамусе.
Новейшие исследования реактивности хвостатого ядра в ответ на непосредственное раздражение в хронических условиях дали очень интересные результаты. При раздражении хвостатого ядра были описаны моторные реакции — поворот головы и круговые движения в противоположную сторону (Хасслер, 8; Бухвальд и Эрвин, 9; Бутхузи, 10). Наш сотрудник Тевзадзе (11) раздражал хвостатое ядро через вживленные электроды и наблю-
|
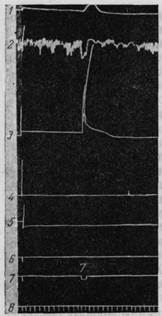
Рис. 192. Двигательная реакция от раздражения хвостатого ядра.
1 — движение головы, 2 — дыхание Регистрация движения конечностей 3 — правая передняя, 4 — левая передняя, 5 — левая задняя, 6 — линия условного сигнала, 7 — безусловное раздражение (стимулятор, 7 в), 8 — время по 2 сек. В ответ на раздражение хвостатого ядра слева наступает поворот головы направо, задержка дыхания и сильное сгибание правой передней ноги (Тевзадзе).
дал определенные внешние реакции. При раздражении среднего и заднего участка наблюдались медленно нарастающее отклонение головы в противоположную сторону и задержка дыхания. После раздражения голова сейчас же принимала среднее положение, а дыхание восстанавливалось постепенно. При раздражении переднего участка головки хвостатого ядра к названным реакциям присоединялось сгибание противоположной передней ноги (рис. 192). Эти наблюдения были сделаны при фиксации животного в камере. Но в условиях свободных движений те же раздражения хвостатого ядра производили не только поворот головы в противоположную сторону, но и вращение животного в сторону поворота головы. При этом нельзя было заметить никаких внешних выражений эмоционального характера. Далее, известно, что электрическое раздражение хвостатого ядра производит торможение тех корковых движений, которые вызываются непосредственно раздражением двигательной зоны большого мозга (Меттлер и др., 12; Фримен и Красно, 13), но не тормозит движения конечностей, вызванные электрическим раздражением кожи (Тевзадзе, 11).
Электрофизиологическими исследованиями (Бутхузи, 14, 15, 16) было показано, что различные области коры прямыми путями связаны с хвостатым ядром. Так, на ненаркотизированных кошках было показано, что при раздражении сенсомоторной, зрительной и ассоциативных областей коры в хвостатом ядре регистрируются вызванные потенциалы с коротким периодом — 2—5 мсек. (рис. 193).
Эти данные о связях различных областей коры с хвостатым ядром подтверждаются морфологическими исследованиями Кармен и др. (17) и Вебстера (18).
С другой стороны, хвостатое ядро имеет эфферентные связи с рядом подкорковых образований: с гипоталамусом, ретикулярной формацией среднего мозга и черной субстанцией (Бутхузи, 19).
Итак, по новейшим исследованиям, базалъные ганглии обладают связями как с корой, так и с другими подкорковыми образованиями и как будто являются передаточными станциями от коры к промежуточному мозгу, а также в центростремительном направлении к коре мозга. При раздражении они производят определенные движения не поведенческого характера и одновременно общее торможение коры и подкорковых образований.
Спрашивается: можно ли образовать условный рефлекс на базе этих двигательных эффектов и каково участие их вообще в индивидуально приобретенной деятельности?
Опыты показали, что все двигательные реакции от раздражения хвостатого ядра могут послужить основой для образования условных рефлексов как на звуки (тон 600 гц) — после 6 сочетаний, так и на свет электрической лампы (75 свечей) — после 16 сочетаний. После образования условного рефлекса условные сигналы вызывали точно такие же движения, как безус-
ловное раздражение (рис. 194). Такой же условный рефлекс образовался на электрическое раздражение коры зрительного анализатора, которое само не давало никаких внешних реакций. После 15 сочетаний с электрическим раздражением хвостатого ядра раздражение зрительной коры в отдельности вызывало тот же самый двигательный рефлекс, как при раздражении хвостатого ядра (Тевзадзе, 11).
Временные связи образовываются между хвостатым ядром и корой и в том случае, если условным сигналом является раздражение хвостатого ядра, которое не вызывает движение, а безусловным рефлексом является движение передней или задней конечности от раздражения соответствующей двигательной зоны. После 40—55 сочетаний раздражение хвостатого ядра вызывало сгибание противоположной конечности (рис. 195).
Заслуживает внимания еще то, что раздражение хвостатого ядра не только может стать условным сигналом к возбуждению корковых движений, но и оказывать угнетающее действие на другие условные рефлексы, например на условное пищевое поведение, выработанное на звук, на условное движение конечности, выработанное на раздражение зрительной коры путем сочетания с раздражением двигательной зоны (Тевзадзе, 11).
Бухвальд и др. (20) показали, что редкое раздражение хвостатого ядра вызывает торможение условного рефлекса нажатия лапы на рычаг для получения пищи. Такие же данные получили Бюзер и сотр. (21). Следует отметить, что при этом торможение условной моторной реакции одинаково хорошо получалось и при раздражении медиальных таламических ядер. Возникновение медленных волн в коре больших полушарий или реакции вовлечения в этих опытах указывает на то, что активируются стрио-таламические механизмы, производящие торможение коры.
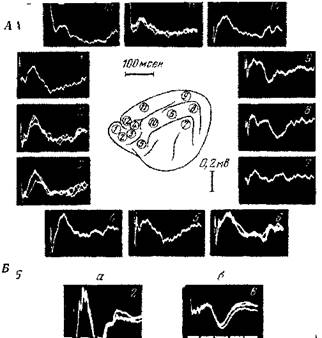
Рис. 193. А — вызванные потенциалы п хвостатом ядре при раздражении различных участков новой коры
Цифры на осциллограммах соответствуют раздражаемым областям коры на схеме мочга
В—ответы в хвостатом ядре при раздражении моторной (а) и зрительной (б) областей коры. Большая развертка — время 5 мсек. (Еутхузи)
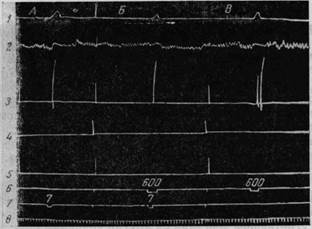
Рис. 194. Условный двигательный рефлекс на правой передней ноге, выработанный на основе раздражения хвостатого ядра
Обозначения кривых и сигнальных линий, как на рис. 192 А — эффект раздражения хвостатого ядра слева поворот головы, остановка дыхания и сгибание правой передней ноги, Б— сочетание условного тона 600 гц, с раздражением хвостатого ядра, В — условный рефлекс на тон 600 гц: поворот головы, задержка дыхания и двукратное сгибание правой передней ноги (Тевзадзе)
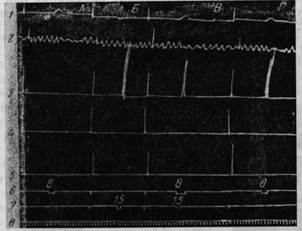
Рис. 195. Условный двигательный рефлекс на правой передней конечности в ответ на условное раздражение хвостатого ядра
Рефлекс вырабатывается на основе раздражения двигательной зоны в левом полушарии А — эффект раздражения хвостатого ядра: поворот головы и ослабление дыхания, Б — эффект раздражения двигательной зоны в левом полушарии учащение дыхания и сгибание правой передней ноги, В — сочетание того и другого раздражения, Г — условное движение правой ноги на раздражение хвостатого ядра после 55 сочетаний Обозначения, как на рис 192 (Тевзадзе).
В острых опытах было показано, что при раздражении хвостатого ядра в коребольших полушарий регистрируется ответ с коротким скрытым периодом (1—2 мсек), и поздняя реакция в виде веретенообразной активности медленных волн (Бутхузи, 16). После повреждения таламических неспецифических ядер ответ с коротким скрытым периодом сохраняется, но исчезает поздняя веретенообразная активность медленных волн (Бутхузи, 16). А из данных Пурпура, Грундфеста и др. (22) известно, что раздражение хвостатого ядра и тогда меняет электрическую активность коры, когда кора изолирована от ствола мозга.
Таким образом, эти данные показывают, что хвостатое ядро может влиять на кору больших полушарий как прямо, так и через таламические неспецифические структуры.
Итак, хвостатое ядро, должно быть, находится с новой корой в таком же взаимодействии, как разные участки коры между собой. Но нервные элементы, образующие временные связи, находятся только в коре в виде ассоциационных и вставочных пирамидных нейронов. Видимо, из корковых временных кругов значительная часть экстрапирамидных путей проходит через ядра, попутно активируя их своими коллатералями.
Дата добавления: 2017-01-13; просмотров: 780;