Общее торможение в коре большого мозга.
Впервые явление общего торможения коры наряду с возбуждением определенных нервных кругов было отмечено при раздражении коры большого мозга в отношении двигательных эффектов, которые вызывались раздражением чувствительных нервов (Беритов и Гедевани, 1). На кошках при барбитуровом наркозе оно вызывалось тетаническим электрическим раздражением любого участка коры. Во время

|
Рис. 58. Общее торможение в коре большого мозга кошки
Регистрируются разгибатели и сгибатели голеностопного сустава: gastrocnemius dex. и sin. (g. d. g. s ), tibialis ant. dex. и sin. (T. d., T. s.,). В опыте А раздражение п. saphenus dex. (S. d.) вызывает сильные сокращения на сгибателях и небольшое сокращение в правом разгибателе. В опыте В то же раздражение нерва происходит во время раздражения (С. s ) ;немоторной зоны в теменной доле того же полушария.
Теперь оно дает ничтожный двигательный эффект.В опыте С через 2 мин. повторяется раздражение нерва и опять сно дает сильно ослабленный двигательный эффект. В опыте D через 4 мин. оно вызывает обычный двигательный эффект (Беритов и Гедевани)
такого раздражения коры тормозились двигательные эффекты, полученные при раздражении чувствительных нервов. Пороги коркового тормозящего раздражения были значительно ниже, чем это нужно для вызова двигательных эффектов при раздражении моторной зоны. Торможение развивалось в первую секунду раздражения. При этом тормозились двигательные эффекты, вызываемые раздражением чувствительных нервов как ипсилатеральной, так и контралатеральной задней конечности (рис. 58). По прекращении коркового раздражения торможение продолжалось в течение нескольких минут, значительно дольше, чем на спинальном препарате, где по прекращении раздражения общее торможение в отношении рефлексов продолжается секунды (Беритов и сотр., 2).
Подобное общее торможение при корковом раздражении получалось в отношении двигательных эффектов, вызываемых раздражением двигательной зоны (рис. 59). Эти корковые эффекты тормозились раздражением как двигательной зоны, так и теменной и затылочной областей (Гедеванишвили, 3).
При раздражении коры мозга в одном участке торможение простирается на всю кору, но заметно сильнее на соответствующее полушарие, чем на другое. При этом тормозятся рефлекторные реакции, вызываемые раздражением как ипсилатералъного двигательного участка коры, так и контралатералъного, как контралатералъного, так и ипсилатералъного нервов. Отсюда следует, что наблюдаемое при корковом раздражении торможение является общим.
В период угнетения корковой деятельности ослабевает и спонтанная электрическая активность коры больших полушарий. Это угнетение также является общим, ибо простирается на оба полушария. Оно наступает как при
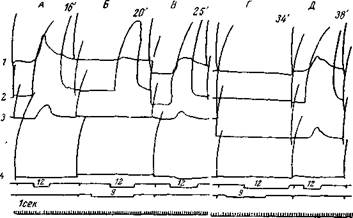
Рис. 59. Общее торможение в коре большого мозга кошки
Регистрируются m quadriceps, m. semitendmosus на обеих конечностях. В опытах А, В и Д раздражается двигательный участок в левом полушарии (верхняя сигнальная линия), при этом наступает ситьное сокращение сгибателя правой стороны (2) и слабое сокращение сгибателя левой стороны и разгибателя правой. В опыте Б и Г это же раздражение во время раздражения немоторной зоны в правом полушдрии (нижняя сигнальная линия) первый раз вызвало лишь ослабленный двигательный эффект, а второй раз не вызвало вообще эффекта. 1 — m. quadriceps dex ,2 — m semitendmosus dex.; 3 — m. semitendmosus sin , 4 — m quadriceps sin (Беритов и Орджоникидзе)
раздражении коры в моторной и немоторной зонах, так и при раздражении чувствительных нервов.
Общее угнетение двигательных реакций при раздражении коры получали также Дюссер де Баренн и Мак Куллоч (4). Но они получали его только при раздражении определенных полос в сенсорной области. Скрытый период этого угнетения измерялся минутами. Авторы пришли к выводу, что оно осуществляется не в коре, а в подкорке. За него ответственны так же такие структуры, как хвостатое ядро, шелуха, бледный шар, зрительный бугор, черная субстанция, мозжечок, т. е. все те структуры, раздражением которых можно обычно вызвать общее торможение. Однако это угнетение наблюдается с большим скрытым периодом (несколько минут) и только при глубокой анестезии (Кларк и сотр., 5).
В наших опытах общее торможение при раздражении коры начинается в первую же секунду и при самом поверхностном наркозе, когда в ответ на раздражение двигательной зоны коры или чувствительного нерва наступает общее движение животного. Кроме того, оно наблюдается при таком подпороговом раздражении двигательной зоны коры, которое не вызывает двигательного эффекта. Все это указывает на то, что наблюдаемое нами общее торможение при раздражении коры действительно вызывается в самой коре.
Электрофизиологические исследования подтверждают, что в опытах с раздражением коры торможение возникает непосредственно в ней. Ройтбак (6), исследуя изменения электрической активности коры при раздражении какого-либо ее участка, обнаружил, что изменение электрической активности происходит во всей коре. При этом, как указывалось выше, потенциал, отводимый на близком расстоянии (не более 10 мм от раздражаемого участка), отрицательный, а в дальних участках он положительный. Отрицательный потенциал наступает здесь после положительного. Это свидетельствует о том, что непосредственное влияние раздражения простирается на ближайшие участки коры (в области отрицательного потенциала), а активация отдельных участков происходит при помощи ассоциационных подкорковых путей. При этом положительный потенциал отражает активность дендритов в IV— JII слоях, а последующий отрицательный — в верхних слоях. И если принять
во внимание, что во время отрицательных медленных колебаний, вызванных раздражением коры, происходит угнетение спонтанной электрической активности, то можно признать, что в вышеприведенных опытах с тетаническим раздражением коры угнетение как двигательных эффектов, так и спонтанной электрической активности было корковое и вызывалось путем активации апикальных дендритов во всей коре. Эта диффузная активация могла произойти как путем передачи импульсов по ассоциационным волокнам через белое подкорковое вещество, так и путем вовлечения таламической восходящей системы, как это бывает при раздражении переднего внутреннего ядра таламуса по Хэнбери и Джасперу (7). В обоих случаях торможение фактически развивается в коре. Весьма возможно, что длительное тормозящее последействие, измеряемое минутами, обусловлено деятельностью кортико-таламокортикальных замкнутых кругов.
Дата добавления: 2017-01-13; просмотров: 784;