Биологическое значение кроссинговера
Хромосомная теория наследственности.
В 1902 г. У.Сэттон в США и Т.Бовери в Германии выдвинули гипотезу о расположении менделевских наследственных факторов (генов) в хромосомах, тем самым, заложив фундамент хромосомной теории наследственности.
Основные положения хромосомной теории наследственности:
1. Гены расположены в хромосомах.
2. Расположение генов линейное.
3. Каждый ген занимает в хромосоме определенный участок (локус).
4. Гены, находящиеся в одной хромосоме образуют группу сцепления.
5. Количество групп сцепления соответствует галоидному набору хромосом.
6. Гены, локализованные в одной хромосоме наследуются сцеплено (совместно).
Сцепление генов впервые было открыто в 1906 году У. Бэтсоном и Р. Пеннетом, обнаружившими совместное наследование генов окраски цветка и формы пыльцы у душистого горошка.
P ♀PpLl X ♂ppll
пурпурные цветки красные цветки
удлиненные пыльца круглая пыльца
F1 P-L- - 4831 (69,5%)
P-ll – 390 (5,6%)
ppL- - 393 (5,6%)
ppll – 1338 (19,3%)
2,8 : 0,2 : 0,2 : 0,8
Объяснение этому явлению дали Т. Морган, А.Стертевант, Г.Меллер и К. Бриджес.
В 1911 – 1918 годах в лаборатории Т.Моргана проводили опыты на мухе дрозофиле Drosophila melanogaster. Она удобна для генетических исследований, так как у нее мало хромосом (4 пары), раннее половое созревание, быстрая смена поколений, большое количество потомков. Для дрозофил легко создать одинаковые условия существования. У дрозофилы при скрещивании анализировали наследование двух пар альтернативных признаков:
В – серое тело
Vg – нормальные крылья
b – черное тело
vg – редуцированные крылья
Первое скрещивание мух проводили по схеме Г.Менделя: Все гибриды были с доминантными признаками – получили единообразие особей первого поколения.
Для выяснения генотипа гибридов провели анализирующее скрещивание – это скрещивание особи c доминантным признаком с рецессивной гомозиготой.
P ♂BbVgvg x ♀bbvgvg

 G BVg bvg
G BVg bvg
Bvg
F BbVgVg : bbvgvg
1:1
родительские типы
50% : 50%
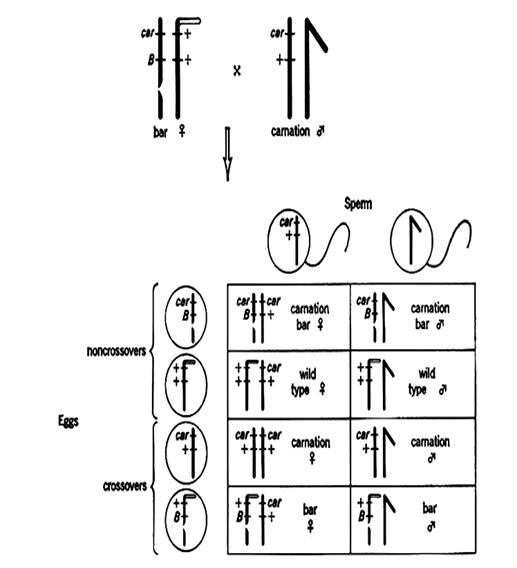
Реципрокное скрещивание
P ♀BbVgvg x ♂ bbvgvg
G BVg
Bvg
Bvg bvg
BVg
 |
F BbVgVg родит. типы
 bbvgvg
bbvgvg
Bbvgvg
bbVgvg рекомб. типы
41,5% : 41,5% : 8,5% : 8,5%
Первое скрещивание позволяет сказать, что самка дрозофилы является дигетерозиготной (BbVv), и что гены цвета тела и длины крыльев находятся в одной паре гомологичных хромосом – они сцеплены с этой парой хромосом. Если бы они были расположены в разных парах гомологичных хромосом, число особей с разными генотипами было бы по 25%.
В реципрокном скрещивании гены обнаруживают частичное сцепление: наряду с родительскими типами (BbVgvg и bbvgvg) в потомстве наблюдаются и рекомбинантные (Bbvgvg и bbVgvg). Появление рекомбинантных типов в потомстве (8,5%+8,5%) Морган объяснил прохождением кроссинговера между гомологичными хромосомами в ходе гаметогенеза. Особи, составляющие по 8,5% образовались в процессе кроссинговера и называются кроссоверными.
Кроссинговер – это обмен гомологичными участками между гомологичными хромосомами. Кроссоверные гаметы – гаметы, содержащие хроматиды, которые прошли кроссинговер. Неизмененные хроматиды входят в состав некроссоверных гамет.
В отличие от независимого комбинирования, когда гены расположены в разных парах хромосом и при скрещивании образуется по 25% особей каждого генотипа, при сцеплении гены расположены в одной паре гомологичных хромосом.
У человека в каждой паре гомологичных хромосом кроссинговер имеет место в среднем в 2-3х точках.
В начале 30-х годов К. Штерн исследовав Х-хромосомы D/ melanogaster, которые имели морфологические различия, получила цитологические доказательства кроссинговера.
Кроссоверные самки содержали в своем кариотипе преобразованные в результате кроссинговера X-хромосомы – длинную палочковидную или двуплечую с короткими плечами
Биологическое значение кроссинговера
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых представляет собой как единый суперген, контролирующий несколько признаков. В то же время, в ходе кроссинговера возникают рекомбинации – т.е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
Эволюционное значение кроссинговера. В результате кроссинговера неблагоприятные аллели, первоначально сцепленные с благоприятными, могут переходить в другую хромосому. Тогда возникают новые гаплотипы, не содержащие неблагоприятных аллелей, и эти неблагоприятные аллели элиминируются из популяции.
Пример. Гаплотип Al оказывается неблагоприятным по сравнению с гаплотипом «дикого типа» (++) за счет наличия летального аллеля l. Поэтому аллель А (благоприятный, нейтральный ил несколько снижающий приспособленность) не может проявиться в фенотипе, поскольку данный гаплотип (Al) содержит летальный аллель l. В результате кроссинговера возникают рекомбинантные гаплотипы A+ и +l. Гаплотип +l элиминируется из популяции, а гаплотип A+ фиксируется (даже в том случае, если аллель А несколько снижает приспособленность его носителей).
Сцепление генов – это совместная передача генов одной хромосомы из поколения в поколение. Сцепление называется полным, если не образуются кроссоверные особи (самец дрозофилы). Если образуются кроссоверные особи (самка дрозофилы), сцепление будет неполным. Кроссинговер происходит не всегда, поэтому кроссоверных особей всегда меньше, чем некроссоверных. Сила сцепления между генами (частота кроссинговера) зависит от расстояния между ними: чем больше расстояние, тем слабее силы сцепления, тем чаще происходит кроссинговер.
1% кроссинговера равен 1 морганиде – единица расстояния между генами, названная в честь Т.Моргана. (В описанном эксперименте общее количество кроссоверных особей – 17% – соответствует расстоянию между генами цвета тела и длины крыльев – 17 морганид).Сцепление может быть аутосомным (группы сцепления аутосом) и гоносомным (группы сцепления половых хромосом).
Используя полученные данные частот рекомбинаций между генами, ученик Моргана Стертевант впервые определил порядок расположения генов в хромосоме: он доказал, что расположение генов в хромосоме должно быть линейным. Им была построена первая генетическая карта X –хромосомы дрозофилы, на которую было нанесено пять генов.
n y – (yellow) – желтое тело;
n w – (white) – белые глаза;
n v – (vermilion) – киноварный цвет глаз;
n m – (miniature) – миниатюрные глаза;
n r – (rudimentary) – рудиментарные крылья.

Таким образом, зная расстояние между генами, можно построить карты хромосом.
Генетическая карта: хромосома представлена в виде прямой, на которой по результатам анализирующего скрещивания условно располагают гены.
Цитологическая карта – точный рисунок или фотография хромосомы. Порядок расположения генов определяют при сравнении результатов анализирующего скрещивания и хромосомных перестроек.
Генетическое картирование осуществляется с применением разных методик.
Генетическое картирование проводится путем генетического анализа - построения генетических карт или карт сцепления.
Генетическое картирование проводится путем параллельных исследования на человеке и на модельных объектах (млекопитающих)
Генетическое картирование проводится с использованием СЕРН-коллекций родословных.
CEPH (Centre d'Etudes du Polymorphysme Humain) - коллекция перевиваемых клеточных культур, полученных от членов семей, многоступенчатые родословные которых насчитывают десятки и сотни индивидуумов.
Цитогенетическое картирование проводят с использованием методов дифференциального окрашивания хромосом, который позволяют идентифицировать при многоцветной окраске (multicolor banding) любой участок хромосомы (бэнд) К основным методам формирования цитогенетических карт относятся также - хромосомный сортинг (проточная цитометрия), микродиссекции и микроклонирование определенных геномных фрагментов и сравнительное генетическое картирование.
Дата добавления: 2016-10-17; просмотров: 2152;