Половой диморфизм в онтогенетической эволюции человека
Половой диморфизм охватывает как самые ранние, так и самые поздние периоды человеческой жизни, не ограничиваясь периодами половозрелости и полового созревания, т. е. относится к постоянным характеристикам онтогенетической эволюции человека, видоизменяющимся лишь по степени интенсивности (усиления или ослабления полового диморфизма).
Независимо от степени интенсивности половой диморфизм является постоянным принципом дифференциации онтогенетической эволюции. Современное естествознание располагает серьезными аргументами в пользу положения, что эта дифференциация не ограничивается только половыми органами и функциями, но охватывает сплошную иерархию «вторичных», «третичных» и еще более отдаленных от половых функций признаков, относящихся к общесоматическим системам и функциям. Переходы между специфическими и общими характеристиками полового диморфизма многообразны и многоступенчаты. Эти характеристики, возможно, относятся ко всем уровням организации – от молекулярного, клеточного и тканевого до организменного.
Можно допустить, учитывая особую роль эндокринной системы и биохимических факторов в определении пола, что обусловленная ими модификация системы связана со всеми основными механизмами регулирования жизненных процессов. Онтогенез человека раскрывается как последовательная смена возрастов, одной из характеристик которых является своеобразный половой метаморфоз: постепенное развертывание и усиление полового диморфизма в периоды роста и созревания, затем стабилизация этого диморфизма, по которой определяется половозрелость, наконец ослабление и угасание полового диморфизма в процессе старения.
Капитальнейшими фактами онтогенетической эволюции являются латентное состояние и постепенное развертывание главнейших феноменов полового созревания, весьма лабильное и многофазное изменение половозрелого состояния, многообразие явлений полового увядания, составляющего один из центральных компонентов старения. Возрастные метаморфозы созревания, зрелости и старения представляют комплексы общесоматических и специфически половых характеристик. Хотя не существует полного параллелизма между общесоматическими и сексуальными (соматоэндокринными) изменениями, с одной стороны, и нервно-психическими, с другой, однако они всегда причинно и функционально взаимосвязаны.
Для понимания интимных взаимосвязей между специфическими (половыми) и общими (соматическими) структурно-динамическими характеристиками организма может оказаться полезной схема системы взаимозависимостей в развивающемся организме, предложенная известным советским биологом акад. И. И. Шмальгаузеном. Исходя из того, что основным элементом высших формообразовательных механизмов является индукционная система из двух взаимодействующих систем, из которых одна выделяется активностью в своем воздействии на другую (индуктор), а вторая – реактивностью, т. е. высвобождением внутренних запасов энергии (реактор), И. И. Шмальгаузен пришел к выводу, что отношения между активирующей и реагирующей тканью составляют один из основных контуров биологического регулирования. По генетическому порядку он является вторым в эмбриогенезе, непосредственно связанным с первым контуром регулирования связей между ядром и плазмой. В свою очередь, второй контур (индуктор–реактор) прямыми и обратными связями включается в обширный третий контур регулирования (эндокринная–нервная система).
Примечательно, что, говоря об индукторе и реакторе, И. И. Шмальгаузен пишет об их условном подразделении, так как «в процессе взаимодействия они постоянно меняются своими местами. Всегда реагирующая часть оказывает и обратное влияние на индуктор, да, кроме того, и сама приобретает значение индуктора для других частей»*. В этом свете получают объяснение взаимосвязи между специфическими и общими функциями целостного организма. Вот что пишет по этому поводу И. И. Шмальгаузен: «Развитие половых признаков контролируется у позвоночных в большей или меньшей мере эндокринной системой... Непосредственная детерминация определяется количественными соотношениями между мужскими и женскими гормонами при установлении пороговых уровней нормальной реактивности тканей. Между всеми органами внутренней секреции имеются сложные взаимодействия, ведущие к регулированию их функций. В случае половых желез выявляется, кроме гипофиза, также влияние щитовидной железы и надпочечников. С другой стороны, на деятельность эндокринной системы, прежде всего гипофиза, влияют и факторы внешней среды. В некоторых случаях установлено и наличие обратной связи между развивающимся вторичнополовым признаком и половыми железами»** (курсив наш. – Б. А.).
* Шмальгаужн И. И. Регуляция формообразования в индивидуальном развитии. – М: Наука, 1964. – С. 123.
** Шмалъгаузен И. И. Регуляция формообразования в индивидуальном развитии. – М: Наука, 1964. – С. 125-126.
Представление о многоконтурной системе биологического регулирования и изменении взаимоотношений между индукторами и реакторами позволяет понять изменяющееся взаимодействие между специфически половыми и общесоматическими явлениями развития, носящее характер саморегулирования. Благодаря многообразным обратным связям от реактора к индуктору, посредством которых замыкаются контуры биологического регулирования, явления полового диморфизма, очевидно, и не могут быть связаны только с метаморфозом половых функций в процессе онтогенетической эволюции.
Не вызывает никакого сомнения, что половая дифференциация онтогенеза человека является эффектом и постоянным условием регулирования всех процессов внутренней среды организма. Но в каком отношении находится эта дифференциация к регулированию процессов взаимодействия организма с внешней средой?
Несомненно, что этот род взаимодействия регулируется непосредственно нервной системой и только через нервную систему проявляется участие эндокринной системы в динамике связей организма со средой обитания. Заметим, кстати, что в своей трехконтурной схеме регулирования процесса формообразования в онтогенезе И. И. Шмальгаузен поместил нервную систему во внешнем контуре, у входа и выхода системы регулирования взаимосвязей организма со средой. Это не значит, конечно, что нервная система не регулирует состояния внутренней среды. Против этого говорят все факты, свидетельствующие о рефлекторной природе гомеостаза. Тем более это не означает, что нервная система, стоящая «у выхода» во внешнюю среду, т. е. организующая деятельность организма в окружающем мире, не испытывает влияния колебаний внутренней среды.
Из общей теории биорегулирования известно, что иерархический, многоуровневый принцип построения рефлекторно-кольцевых механизмов регулирования выполняет с высокой надежностью задачи взаимоувязывания внешней и внутренней среды организма. Благодаря центроэнцефалической системе большие полушария головного мозга многообразно используют для регулирования внешней деятельности, поведения мощные энергетические и информационные потоки, генерируемые всеми системами и тканями организма.
Вероятно, что именно через нервную систему, а не непосредственно через эндокринные аппараты на большие полушария могут влиять специфически половые характеристики организма*. Вместе с тем их влияние на центроэнцефалическую систему и через нее на подкорковые аппараты больших полушарий, несомненно, всегда значительно. Кора больших полушарий, непосредственно являющаяся субстратом временных связей организма с внешней средой, испытывает «снизу», из всех нижележащих отделов центральной нервной системы, тонизирующее, активирующее воздействие; но еще более важно то, что в соответствии с объективными условиями внешней среды большие полушария непрерывно реорганизуют, а вместе с тем и усиливают устойчивость внутренней среды организма через тормозящие воздействия на нижележащие отделы центральной нервной системы (ц. н. с.).
* Возможно, что этот механизм определяет общность «ростовой, сексуальной и интеллектуальной акселерации», их параллелизм в подростковом возрасте. Ряд психофизических корреляций в этом возрасте описан в кн.: Гримм Г. Основы конституциональной биологии и антропометрии. – М.: Медицина, 1967.
Взаимопроникновение общесоматических и половых характеристик в динамике основных функций жизнедеятельности человека всегда учитывается в диагностической практике. Начиная с сердечного ритма (у женщин, как известно, более частого сравнительно с мужчинами) и кончая характеристиками обмена веществ, все больше уясняется фактор полового диморфизма в жизнедеятельности человека.
Энергетическая потребность (в калориях) здоровых людей на 1 м2 поверхности тела в 1 ч, по данным Дю-Буа и Оуба*, равна:
* Приведено по кн.: Павелъски С., Заводски 3. Физиологические константы в клинике внутренних болезней. - М.: Медицина, 1964. - С. 228.
| Возраст, лет | Мужчины | Женщины |
| 16-18 | 43,0 | 40,0 |
| 18-20 | 41,0 | 38,0 |
| 20-30 | 39,5 | 37,0 |
| 30-40 | 39,5 | 36,0 |
| 40-50 | 38,5 | 36,0 |
| 50-60 | 37,5 | 35,0 |
| 60-70 | 36,5 | 34,0 |
| 70-80 | 33,5 | 33,0 |
Энергетическая потребность стабилизируется у мужчин в период 20-40 лет, а у женщин – в 30-50 лет. Любопытно, что половой диморфизм сохраняется и в старости, когда непосредственное влияние половых функций все более ослабляется. Даже в том случае, если вес тела идентичен у мужчин и женщин одного и того же возраста, все равно «должный калораж» различен, как это вычислено Харрисом и Бенедиктом* по отношению к основному обмену.
* Там же. - С. 229.
Сравнительно-возрастная физиология накопила необычайно много самых разнообразных данных, смысл которых еще недостаточно ясен. Вот, например, данные о возрастных изменениях основного обмена у здоровых мужчин и женщин.
По сводке Н. Шока, приводимой Бурльером, основной обмен (определяемый в 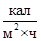 ) у мужчин изменяется весьма значительно: в 20 лет величина основного обмена равна 42,52 ± 1,60, а в 60 лет снижается до 35,22 ± 1,71. Между тем у женщин эта величина более постоянна, снижение ее с возрастом меньше: в 20 лет величина основного обмена – 36,73 ± 2,65, а в 60 лет – 34,68 + 2,46. Таким образом, существует половое различие в тенденции возрастных изменений основного обмена.
) у мужчин изменяется весьма значительно: в 20 лет величина основного обмена равна 42,52 ± 1,60, а в 60 лет снижается до 35,22 ± 1,71. Между тем у женщин эта величина более постоянна, снижение ее с возрастом меньше: в 20 лет величина основного обмена – 36,73 ± 2,65, а в 60 лет – 34,68 + 2,46. Таким образом, существует половое различие в тенденции возрастных изменений основного обмена.
Наибольшее расхождение в величинах основного обмена у мужчин и женщин можно отметить в 20 лет (у мужчин – 42,52, у женщин – 36,73), а наименьшее – в 60 лет (у мужчин – 35,22, у женщин – 34,68). Интересно отметить, что и в возрастной эволюции секреции и состава желудочного сока (взятого после пробного завтрака и измеряемого в миллилитрах) обнаруживается сходная тенденция.
У мужчин количество желудочного сока изменяется со 113,5 мл (в 20–29 лет) до 96,3 мл (в 50-59 лет). За этот же период времени у женщин количество желудочного сока изменяется с 99,0 мл (в 20-29 лет) до 93,7 мл (в 50–59 лет), т. е. обнаруживается значительно большая стабильность количества желудочного сока, а возможно, и всех функций пищевого обмена.
Поразительны в этом отношении данные Де-Смета, Шока и Вандер-Вельде о возрастных изменениях кислотности желудочного сока. Свободная кислотная у мужчин в 20-29 лет равна 47,0 мл (1 мл 0,1 н. раствора NaOH на 100 мл сока), а в 50-59 лет – 42,6. У женщин же свободная кислотная, равная 33,0 мл, остается без изменений в три возрастных периода (20-29, 30-39 и 40-49 лет) и только в период 50-59 лет незначительно меняется, достигая 33,3 мл. Что касается общей кислотности желудочного сока, то у мужчин она изменяется за 40 лет с 62,5 мл (в 20 лет) до 57 (в 60 лет), а у женщин эта величина имеет значение абсолютной константы для всех возрастных периодов*.
* Бурльер Ф. Старение и старость. - М: ИЛ, 1962. - С. 24.
Дата добавления: 2016-09-20; просмотров: 1117;