Трофическая структура биоценозов. Основная функция биоценозов - поддержание круговорота веществ в биосфере - базируется на пищевых взаимоотношениях видов
Основная функция биоценозов - поддержание круговорота веществ в биосфере - базируется на пищевых взаимоотношениях видов. Именно на этой основе органические вещества, синтезированные автотрофными организмами, претерпевают многократные химические трансформации и в конечном итоге возвращаются в среду в виде неорганических продуктов жизнедеятельности, вновь вовлекаемых в круговорот. Поэтому при всем многообразии видов, входящих в состав различных сообществ, каждый биоценоз с необходимостью включает представителей всех трех принципиальных экологических групп организмов - продуцентов, консументов и редуцентов. Полночленность трофической структуры биоценозов - аксиома биоценологии (в гидробиологии иногда принято говорить о неполночленных биоценозах, существующих за счет энергии привносимого извне органического вещества, но по сути дела такие системы не самостоятельны и не обладают способностью к самоподдержанию).
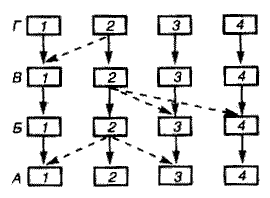
В конкретных биоценозах продуценты, консументы и редуценты представлены популяциями многих видов, состав которых специфичен для каждого отдельного сообщества. Функционально же все виды распределяются на несколько групп в зависимости от их места в общей системе круговорота веществ и потока энергии. Равнозначные в этом смысле виды образуют определенный трофический уровень, а взаимоотношения между видами разных уровней - систему цепей питания. Совокупность трофических цепей в их конкретном выражении, включающем прямые и косвенные взаимоотношения составляющих их видов, формирует целостную трофическую структуру биоценоза (рис. 13.1).
Трофические уровни. Группа видов-продуцентов образует уровень первичной продукции, на котором утилизируется внешняя энергия и создается масса органического вещества. Первичные продуценты - основа трофической структуры и всего существования биоценоза.
|
| Рис, 13.1. Простейшая схема трофической структуры биоценоза (по И.А. Шилову, 1985) А-Г - трофические уровни. А - продуценты, Б - консументы I порядка (фитофаги), В - консументы II порядка, Г - консументы III порядка; 1 - 4-конкретные виды данного трофического уровня; одинаковыми цифрами обозначены отдельные трофические цепи; сплошными стрелками - прямые (трофические) связи, пунктирными стрелками-косвенные (конкурентные) связи. |
Составлен этот уровень растениями (кроме редких бесхлорофилльных форм) и фо-тоавтотрофными прокариотами; в особых случаях в качестве первичных продуцентов выступают бактерии-хемосинтетики. В гл. 1 уже упоминались своеобразные глубоководные экосистемы, привязанные к разломам земной коры и основывающиеся на деятельности хемоавтотрофных бактерий.
Биомасса органического вещества синтезированного автотрофами, определяется как первичная продукция, а скорость ее формирования - биологическая продуктивность экосистемы. Продуктивность выражается количеством биомассы, синтезируемой за единицу времени (или энергетическим эквивалентом). Общая сумма биомассы рассматривается в этом случае как валовая продукция, а та ее часть, которая определяет прирост,- как чистая продукция. Разница между валовой и чистой продукцией определяется затратами энергии на жизнедеятельность ("затраты на дыхание"}, которые, например, в умеренном климате могут составлять до 40-70 % валовой продукции.
Накопленная в виде биомассы организмов-автотрофов чистая первичная продукция служит источником питания для представителей следующих трофических уровней. Потребители первичной продукции - консументы - образуют несколько (обычно не более 3-4) трофических уровней.
Консументы I порядка. Этот трофический уровень составлен непосредственными потребителями первичной продукции. В наиболее типичных случаях, когда последняя создается фотоавтотрофами, это растительноядные животные (фитофаги). Виды и экологические формы, представляющие этот уровень, весьма разнообразны и приспособлены к питанию разными видами растительного корма. В связи с тем, что растения обычно прикреплены к субстрату, а ткани их часто очень прочны, у многих фитофагов эволюционно сформировался грызущий тип ротового аппарата и различного рода приспособления к измельчению, перетиранию пищи. Это зубные системы грызущего и перетирающего типа у различных растительноядных млекопитающих, мускульный желудок птиц, особенно хорошо выраженный у зерноядных, и т. п. У растительноядных карповых рыб над глоточными зубами, на нижней поверхности мозгового черепа имеется образование - "жерновок", имеющий ороговевшую поверхность; сочетание этих структур определяет возможность перемалывания твердой пищи. Грызущий ротовой аппарат свойствен многим насекомым и др.
Некоторые животные приспособлены к питанию соком растений или нектаром цветков. Эта пища богата высококалорийными, легкоусвояемыми веществами. Ротовой аппарат у питающихся таким образом видов устроен в виде трубочки, с помощью которой всасывается жидкая пища.
Приспособления к питанию растениями обнаруживаются и на физиологическом уровне. Особенно выражены они у животных, питавшихся грубыми тканями вегетативных частей растений, содержащими большое количество клетчатки. В организме большинства животных нe продуцируются целлюлозолитические ферменты, а расщепление клетчатки осуществляется симбиотическими бактериями (и некоторыми простейшими кишечного тракта).
Среди млекопитающих переработка клетчатки особенно специализирована у жвачных копытных, желудок которых имеет сложное строение (рис. 13.2). Наиболее объемистый его отдел - рубец -служит вместилищем, в котором проглоченный корм перемешивается со слюной и подвергается воздействию ферментов симбиотических бактерий и простейших. Продукты бактериологического сбраживания клетчатки (в основном летучие жирные кислоты - уксусная, янтарная, масляная) всасываются здесь же, в рубце. Оставшаяся часть растительной массы после вторичного пережевывания подвергается действию желудочного сока в другом отделе желудка-сычуге-и алее следует обычным путем через кишечник, последовательно обрабатываемая различными ферментами. При таком типе питания в кишечник наряду с растительной массой попадает большое количество бактерий и простейших, клетки которых служат источником белкового питания; кроме того, некоторые бактерии в рубце способны синтезировать белок из аммонийных солей и мочевины.
У других млекопитающих основным местом переработки клетчатки вляется толстая кишка, особенно ее слепой вырост. Химизм этих процессов сходен с пищеварением в рубце. У зайцеобразных и многих грызунов в связи с таким типом пищеварения выделяется два типа помета: первичный, который животные заглатывают, и вторичный более сухой, который выводится из организма. Поедание первичного помета способствует утилизации содержащегося в нем белка (включая и бактериальные клетки), а может быть, и "подсеву" регулярно теряемой с экскрементами кишечной флоры. Аналогичным образом g области толстого кишечника и его слепых выростов идет переваривание клетчатки у питающихся растительными тканями птиц (куриные, гусеобразные, врановые и др.).
Переваривание клетчатки с участием бактерий и простейших цщ-роко распространено и среди беспозвоночных животных. У низцщх термитов эта функция осуществляется главным образом с помощью простейших (жгутиковые), у высших термитов и других групп большее значение имеют бактерии. Впрочем, среди беспозвоночных имеются и формы, продуцирующие собственные ферменты, расщепляющие клетчатку. Так, примитивные насекомые чешуйницы Ctenolepisma lineata переваривают ее даже в условиях полного освобождения кишечника от бактерий (R. Lasker, A. Giese, 1956). Продукция целлюлозолитических ферментов, возможно, свойственна моллюску Teredo ("корабельный червь") и некоторым другим животным, питающимся древесиной.
Консументы частично используют пищу для обеспечения жизненных процессов ("затраты на дыхание"), а частично строят на ее основе собственное тело, осуществляя таким образом первый, принципиальный этап трансформации органического вещества, синтезированного продуцентами. Процесс создания и накопления биомассы на уровне консументов обозначается как вторичная продукция.
Консументы II порядка. Этот уровень объединяет животных с плотоядным типом питания (зоофаги). Обычно в этой группе рассматривают всех хищников (понятие "хищничество" в данном случае используется в широком экологическом смысле как плотоядный тип питания, при котором животное-зоофаг, нападает на жертву и убивает ее), поскольку их специфические черты практически не зависят от того, является ли жертва фитофагом, или плотоядна. Но, строго говоря, консументами II порядка следует считать только хищников, питающихся растительноядными животными и соответственно представляющих второй этап трансформации органического вещества в цепях питания. Химические вещества, из которых строятся ткани животного организма, довольно однородны, поэтому трансформация при переходе с одного уровня консументов на другой не имеет столь принципиального характера, как преобразование растительных тканей в животные.
При более тщательном подходе уровень консументов II порядка следует разделять на подуровни соответственно направлению поток вещетва и энергии. Например, в трофической цепи "злаки - кузнечики -лягушки - змеи - орлы" лягушки, змеи и орлы составляют последовательные подуровни консументов II порядка.
Зоофаги характеризуются своими специфическими приспособлениями к характеру питания. Например, их ротовой аппарат часто приспособлен к схватыванию и удержанию живой добычи. При питанииl животными, имеющими плотные защитные покровы, развиваются приспособления для их разрушения. Таковы специализированные зубы ("терка") скатов, раздавливающих панцири иглокожих и раковины моллюсков, или снабженные округлыми бугорками широкие коренные зубы каланов Enhydra lutris, питающихся преимущественно морскими ежами. Вороны и крупные чайки иногда, схватив твердую добычу (моллюски, крабы и т. п.), взлетают и с высоты бросают ее на землю; этот прием они повторяют до тех пор, пока раковина или панцирь не расколется. Возможно, что аналогичным образом поступают некоторые хищные птицы с черепахами (стервятники) или с костями крупных животных (бородач).
На физиологическом уровне адаптации зоофагов выражаются прежде всего в специфичности действия ферментов, "настроенных" на переваривание пищи животного происхождения. В опытах с собаками показано, что такая "настройка" может меняться: предварительное кормление растительными белками (хлеб) усиливает активность пепсина к этим белкам и уменьшает перевариваемость мяса (А.М. Уголев, 1958).
У некоторых насекомоядных млекопитающих и птиц в желудке обнаруживаются ферменты хитиназа и хитобиаза, гидролизующие хитин - сложный азотсодержащий углевод. Хитиназа найдена также в секрете желудочных желез и в панкреатическом соке ряда видов амфибий и рептилий. Обнаружен этот фермент и у водных позвоночных (круглоротые, оба класса рыб), особенно у видов, корм которых содержит много хитина. Происхождение хитиназы у рыб не совсем ясно: есть предположение, что она продуцируется симбиотическими бактериями (Т. Goodrich, R. Morita, 1977). По другим данным активность фермента в слизистой желудка выше, чем в его содержимом, что свидетельствует о секреции его пищеварительными железами. Хити-нолитические ферменты известны и у ряда беспозвоночных (дождевые черви, некоторые ракообразные и моллюски); у многих из них про-дукция хитиназы связана с деятельностью бактерий-симбионтов.
Известен и ряд специфических приспособлений к питанию. Например, хищные моллюски, питающиеся представителями того же ^па, "просверливают" их раковины с помощью концентрированных минеральных кислот, продуцируемых специальными железами. Укровососущих летучих мышей - вампиров - в секрете слюнных желез содержатся антикоагулянты, препятствующие свертыванию крови и тем способствующие длительному питанию. Выделение антикоагулян, тов свойственно также пиявкам и большому числу кровососущих членистоногих.
Консументы III порядка. Выделение этого уровня в достаточной степени условно. Обычно сюда тоже относят животных с плотоядным типом питания, чаще всего имея в виду паразитов животных и "сверхпаразитов", хозяева которых сами ведут паразитический Образ жизни. Явление сверхпаразитизма побуждает и этот трофический уровень делить на подуровни. Но строго говоря, сюда относятся все животные, поедающие плотоядных животных, тогда как паразиты растительноядных форм должны рассматриваться как консументы 2 порядка.
Паразитизм - сложная и многообразная форма взаимоотношений, которая подробнее будет рассмотрена ниже (см. гл. 14). В самец общем виде паразитизм отличается от хищничества тем, что паразит не. убивает свою "жертву" (хозяина), а длительно питается на живом объекте; Гибель хозяев происходит лишь при массовом и длительном заражении и в большинстве случаев невыгодна для паразита.
Деление биоценоза на трофические уровни представляет собой лишь общую схему. Действительные формы взаимоотношений сложнее. Например, существует много видов со смешанным питанием; такие виды могут одновременно относиться к разным трофическим уровням.
Переход биомассы с нижележащего трофического уровня на вышележащий связан с потерями вещества и энергии. В среднем считается, что лишь порядка 10 % биомассы и связанной в ней энергии переходит с каждого уровня на следующий. В силу этого суммарная биомасса, продукция и энергия, а часто и численность особей прогрессивно уменьшаются по мере восхождения по трофическим уровням. Эта закономерность сформулирована Ч. Элтоном (Ch. Eiton, 1927) в виде правила экологических пирамид (рис. 13.3) и выступает как главный ограничитель длины пищевых цепей.
Трофические цепи и сети питания. Прямые пищевые связи типа "растение - фитофаг - хищник - паразит" объединяют виды в цепи питания или трофические цепи (рис. 13.1), члены которых связаны между собой сложными адаптациями, обеспечивающими устойчивое существование каждой видовой популяции.
Уже отмечалось, что каждый трофический уровень составлен не одним, а многими конкретными видами. Видовое разнообразие биоценотических систем - не случайное явление. Благодаря видоспецифичности питания увеличение числа видов в биоценозе определяет более полное использование ресурсов на каждом трофическом уровне. Это обстоятельство прямо связано с повышением полноты биогенного круговорота веществ.
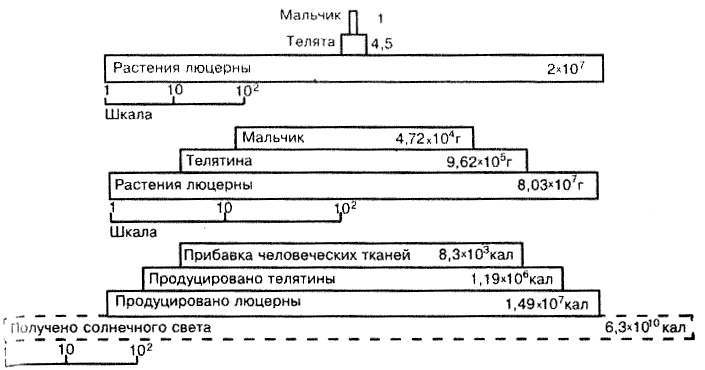
Рис. 13.3. Пример простой экологической пирамиды (по Ю. Одуму, 1975): вверху --пирамида чисел, в середине - биомассы, внизу - энергии. Данные приведены И расчете на 4 га за 1 год
Кроме того, увеличение видового разнообразия выступает как "гарантийный механизм", обеспечивающий надежность круговорота веществ как главной функции экосистем. Суть этого механизма заключается в том, что монофагия - питание лишь единственным видом пищи - встречается в природе редко; немногочисленны и олигофаги, набор объектов питания которых включает небольшое число экологически сходных видов. Большинство животных использует в пищу более или менее широкий набор кормовых объектов. В результате помимо прямых "вертикальных" пищевых связей возникают боковые, объединяющие потоки вещества и энергии двух и более пищевых цепей. Простейший пример таких связей приведен на рис. 13.1. Таким путем формируются пищевые (трофические) сети, в которых множественность Цепей питания выступает как приспособление к устойчивому существованию экосистемы в целом: "дублирование" потоков вещества и энергии по большому числу параллельных трофических цепей поддерживает непрерывность круговорота при всегда вероятных нарушениях отдельных звеньев пищевых цепочек.
Таким образом, многочисленность и разнообразие видового состава биоценоза выступает как важный механизм поддержания его целостности и функциональной устойчивости. Именно на этом основывается актуальность проблемы биологического разнообразия, разрабатываемой в настоящее время на уровне международной комплексной программы.
То обстоятельство, что в пищевых сетях обычны "горизонтальные" связи, объединяет животных одного трофического уровня наличием общих объектов питания. Это означает возникновение пищевой конкуренции между различными видами в составе сообщества. Сильная пищевая конкуренция невыгодна для биоценоза в целом; в процессе эволюционного становления экосистем либо часть видов вытеснялась из состава сообщества, либо формировались межвидовые отношения, ослабляющие силу конкуренции.
Возможности ослабления пищевой конкуренции в значительной мере определяются такими ее показателями, как объем и напряженность (А.А. Шорыгин, 1952). Объем конкуренции определяется числом видов пищи, общих для конкурентов. В этом аспекте ослабление конкуренции может идти через расширение пищевого спектра одного или нескольких конкурирующих видов: увеличение набора кормовых объектов ведет к снижению доли их, совпадающей с пищевым спектром другого вида, т. е. к уменьшению относительного объема конкуренции. Расхождение в питании возможно и на базе совпадающих объектов питания, если отличаются предпочитаемые корма формально конкурирующих видов; фактически это также уменьшает объем конкуренции. Наиболее эффективный путь выхода из конкуренции через уменьшение ее объема -высокая специализация в питании, приводящая к расхождению кормовых спектров.
Напряженность конкуренции определяется соотношением потребности в данном виде корма для видов-конкурентов и ее обилием в Природе. Например, очень важное место в питании околоводных Грызунов (бобр, ондатра, водяная полевка) занимают тростник и осоки. Но эти виды растений представлены в природе сплошными зарослями с высокой биомассой и продуктивностью. Поэтому практически обеспечиваются запросы всех обитающих в таких условиях видов, и конкуренция их принимает формальный характер и не приводит к реальным негативным взаимоотношениям. Подобным же образом практически не возникает конкуренция между различными видами чаек и чистиковых, огромными стаями питающихся на массовых скоплениях криля или на косяках рыб. В случае же совпадения лимитированных по биомассе кормовых ресурсов напряженность конкуренции резко возрастает и может стать причиной вытеснения менее конкурентоспособных видов из состава сообщества.
Соотношение объема и напряженности определяет общую силу конкуренции; этот параметр лежит в основе конкретного проявления различных форм взаимоотношений между конкурирующими видами. Подробнее эти отношения рассматриваются в гл. 14.
При ослабленной силе конкуренции система "горизонтальных" связей выгодна для биоценоза: как уже говорилось, на ее основе увеличивается надежность функционирования экосистем; сложная комбинация прямых и косвенных трофических связей объединяет все виды биоценоза в единое функциональное целое.
Цепи разложения. Все рассмотренные выше процессы связаны с синтезом и трансформацией органического вещества в трофических сетях и характеризуют собой так называемые цепи выедания или "пастбищные цепи". Процессы поэтапной деструкции и минерализации органических веществ обычно выводятся в отдельный блок трофической структуры, называемый цепями разложения (или детритными цепями).
Вычленение детритных цепей связано прежде всего с тем, что минерализация органики практически идет на всех трофических уровнях: и растения и животные в процессе метаболизма редуцируют органическое вещество до диоксида углерода и воды; эти продукты выводятся в окружающую среду так же, как и минеральные соли (последнее особенно свойственно животным). Детритные цепи начинаются с разложения мертвой органики особыми группами консументов - сапрофагами. Животные-сапрофаги механически, а отчасти и химически разрушают мертвое органическое вещество, подготавливая его к воздействию редуцентов. В наземных экосистемах этот процесс сосредоточен преимущественно в подстилке и в почве.
Наиболее активное участие в разложении мертвого органического вещества принимают почвенные беспозвоночные животные (членистоногие, черви) и микроорганизмы. Процесс деструкции идет последовательно, "волны" сапрофагов сменяют друг друга в соответствии с видоспецифичным типом питания (рис. 13.4). Крупные сапрофаги (например, насекомые) лишь механически разрушают мертвые ткани они не являются собственно редуцентами, но готовят субстрат ддд организмов (прежде всего бактерий и грибов), осуществляющих процессы минерализации. Кроме того, бактерии-симбионты способствуют превращению сложных органических веществ в более простые, как бы "встраиваясь" в уровень консументов.
Сообщества организмов-сапрофагов отличаются относительно малой жесткостью организации: в их формировании большое значение имеют стохастические процессы, отдельные виды легко взаимозаменяемы, велика роль внешних факторов и конкурентного исключения (Н.М. Чернова, НА. Кузнецова, 1986).
Таким образом, на уровне консументов происходит разделение потока органического вещества по разным группам потребителей: живое органическое вещество следует по цепям выедания, а мертвое - по цепям разложения. В наземных биоценозах цепи разложения имеют очень большое значение в процессе биологического круговорота; в них перерабатывается до 90 % прироста биомассы растений, попадающей в эти цепи в виде опада. В водных экосистемах большая часть вещества и энергии включается в пастбищные цепи.
Дата добавления: 2016-08-08; просмотров: 1434;