ВЛИЯНИЕ ИСТОРИИ НАУЧЕНИЯ НА СТРУКТУРУ ОПЫТА И ОРГАНИЗАЦИЮ МОЗГОВОЙ АКТИВНОСТИ
Субъективный континуум, как и континуум поведения, представляет собой линейную последовательность сменяющих друг друга состояний, которые соответствуют актам поведения. Эти смены одних наборов функциональных систем на другие связаны с достижением результатов одних актов и возможностью реализации следующих актов поведения. В силу того, что субъект выступает в качестве активного компонента соотношений организма со средой (см. гл. 14), переход к реализации следующей в последовательности функциональной системы происходит за счет ее извлечения из памяти под воздействием внутренних факторов. Факторы, определяющие разные состояния систем и смену реализуемых систем в континууме поведения, были названы межсистемными отношениями.
В соответствии с включенностью конкретного акта в жестко фиксированную последовательность, такую, как, например, врожденные стереотипные формы поведения или аппетентная стадия пищедобывательного цикла, эти отношения могут иметь жестко детерминированный характер. В то же время (как в случае поисковой активности), эти отношения не детерминированы и выбор происходит среди слабо связанных в памяти функциональных систем. Но и жесткая детерминация отношений двух последовательных актов внутри цикла не является однозначной – всегда имеется возможность альтернативы, т.е. перехода к каким-либо другим актам поведения. Подобная возможность реализуется, например, при насыщении и переходе от пищедобывательного поведения к комфортному (отдыху) либо при переходе к ориентировочному поведению в связи с изменениями в среде.
Можно утверждать, что смена актов происходит в результате выбора из ряда альтернатив с разной вероятностью в поведении. Наблюдение с этих позиций за формированием умения играть в многоклеточные «крестики-нолики» позволило создать модель структуры памяти и межсистемных отношений, хорошо описывающую выбор хода в конкретной игре [Александров и др., 1997].
Исследование нейрональной активности в поведении показало, что, наряду с ярко выраженной специфической активацией специализированных нейронов в специфическом поведенческом акте, в их активности имеется дополнительная структура. Эта структурированность неспецифической активности (т.е. активности при осуществлении других актов поведения) проявляется в наличии слабых активации и тормозных пауз, сопровождающих реализацию других актов поведения. Связь таких изменений с конкретным актом имеет, как правило, вероятностный, но достаточно устойчивый (по статистическим критериям) характер.
Исходя из принципа системе-специфичности нейрона и его специализации относительно элемента индивидуального опыта, было выдвинуто положение о разных состояниях, или степенях актуализации элемента опыта в реальном континууме поведения. Нейрофизиологическим выражением степени актуализации специфической функциональной системы при подобном подходе является текущая частота активности специализированного нейрона, варьирующая от нулевых значений при исключении системы из состояния субъекта поведения до максимума при реализации соответствующего акта поведения.
Таким образом, на основе анализа нейрональной активности был сделан вывод об актуализации множества функциональных систем при осуществлении конкретного акта поведения. Обнаружение определенной структуры актуализации конкретной функциональной системы в поведении позволило использовать регистрацию активности нейронов известной специализации для изучения отношений конкретного элемента опыта с другими элементами.
В качестве метода выявления межсистемных отношений используется построение паттернов активности специализированных нейронов во всех актах анализируемого поведения и распределения вероятности активации нейрона в конкретном акте поведения. За активацию принимается не менее чем 1,5-кратное превышение «фоновой» частоты активности. Эти формы представления нейрональной активности иллюстрируются на рис. 15.1 в сравнении с распространенным методом представления активности в виде растров. Особенно яркие результаты дало использование данного метода для решения проблемы отражения истории научения в организации межсистемных отношений.
В силу фиксации этапов научения в виде элементов субъективного опыта история научения индивида является содержанием его опыта. Однако кроме состава элементов опыта, история его формирования может (наряду с логикой поведения) определять отношения систем.
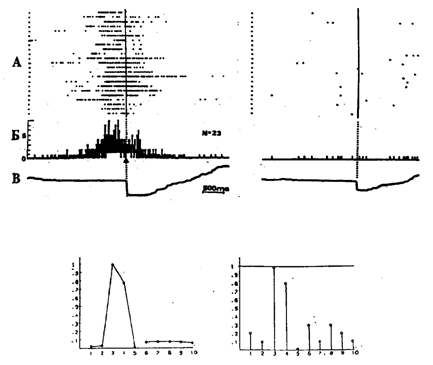
Рис. 15.1. Разные способы представления активности нейрона
А – растры активности нейрона лимбической коры; Б – гистограмма; В – усредненная отметка нажатия педали (слева–левой, справа–правой), относительно начала которой совмещены растры и построены гистограммы. Внизу: графическое представление активности того же нейрона. На левом графике – картина распределения средних частот (паттерн) активности нейрона в актах циклического поведения. По оси абсцисс – номера актов, по оси ординат – нормированная средняя частота активности. На правом графике – вероятности наличия активации в тех же 10 актах. По оси абсцисс – номера актов, по оси ординат – вероятность активации нейрона в соответствующем акте
Было проведено специальное исследование активности нейронов лимбической коры кроликов в пищедобывательном поведении, сформированном с применением разных стратегий обучения. На рис. 15.2 схематически представлены использовавшиеся разные стратегии обучения внешне одному и тому же циклическому пишедобывательному поведению. Разница между стратегиями состояла в последовательности формирования отдельных блоков поведения – поведения у кормушки и поведения у педали – на двух сторонах экспериментальной клетки. Формирование отдельных поведенческих актов производилось так же, как было изложено ранее. На рисунке последовательности этапов научения показаны цепочками стрелок.
Для выявления сформированных межсистемных отношений была зарегистрирована активность нейронов лимбической коры в дефинитивном поведении обученных таким образом животных. Из всей совокупности зарегистрированных нейронов для анализа были отобраны клетки, специализированные относительно сформированных обучением в экспериментальной клетке поведенческих актов.
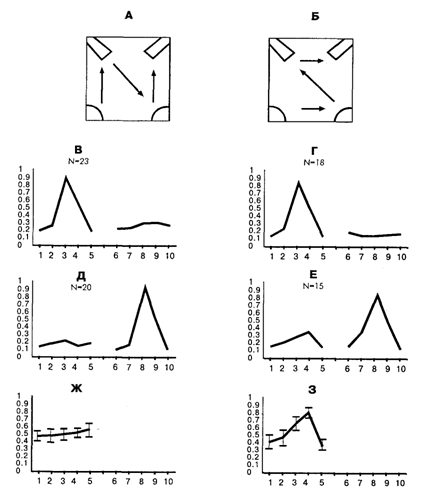
Рис. 15.2 . Усредненные паттерны активности нейронов, специализированных относительно подхода и нажатия педали в зависимости от времени формирования и стратегии обучения
А, Б – схемы применявшихся стратегий обучения (в нижних углах – кормушки, в верхних – педали); В–З: усредненные паттерны нейрональной активности. По оси абсцисс – номера поведенческих актов; по оси ординат – нормализованная по максимальной частота активности; В, Г – усредненные паттерны активности нейронов, специализированных относительно оперирования с первой по порядку обучения педалью; Д, Е– усредненные паттерны активности нейронов, специализированных относительно оперирования со второй по порядку обучения педалью; Ж, 3 – паттерны (средняя ± стандартная ошибка) неспецифической активности нейронов, представленных на частях Д и Е, но нормализованной по максимуму неспецифической активности. Под схемой конкретной стратегии расположены паттерны активности нейронов, которые зарегистрированы у кроликов, обученных по этой стратегии.
Критерием специализации служило наличие активации нейрона в соответствующем акте во всех случаях его реализации. Как было показано А. Г. Горкиным и Д. Г. Шевченко [1990], этот формальный критерий позволяет достаточно надежно выделять специфическую связь активности нейрона с конкретным актом поведения. Для анализа неспецифической активности были построены паттерны активности каждого из исследовавшихся нейронов во всех актах пищедобывательного поведения. Паттерн активности представляет собой распределение средних частот импульсации нейрона в выделенных актах поведения. В поведении на одной стороне клетки было выделено 5 актов: поворот головы к педали, перенос лап к педали, нажатие педали, перенос лап к кормушке и захват пищи в кормушке. Всего на двух сторонах клетки было выделено 10 актов. Усредненные паттерны активности нейронов, специализированных относительно подхода к педали, представлены на рис. 15.2 под схемами соответствующих стратегий обучения. Так как кролик подходил к двум педалям, и паттернов – два для каждой из стратегий. Один из них показывает распределение активности нейронов, специализированных относительно подхода к первой по порядку обучения педали, а другой – относительно второй. Легко заметить, что паттерны у кроликов, обученных по стратегии «Б», различаются для первой и второй педали, в то время как для животных, обученных по стратегии «А», – не различаются. Сходные данные были получены для другой группы специализаций – нейронов, связанных с подходом к кормушке [Горкин, Шевченко, 1995].
Данные, полученные в этой работе, показали, что для систем, не связанных логикой последовательного поведения в одну группировку, их взаимные отношения зависят от истории формирования поведения. Эта зависимость проявляется в виде повышенной степени актуализации второй из двух последовательно формировавшихся систем при реализации первой. В этом исследовании были также выявлены отношения систем, основанные на факторах сходства движений и целей соответствующих поведенческих актов.
В связи с тем, что актуализация функциональных систем, оказалась чувствительной к действию множества факторов, оказывается возможным рассмотреть с позиций системной психофизиологии огромный объем экспериментальных данных, полученных в нейрофизиологии обучения (см. например [Рабинович, 1975; Котляр, 1989]). В основном эти данные касаются пластичности нейрональных ответов на стимулы при выработке условных рефлексов. Сразу стоит обратить внимание на то, что достоверные изменения частоты постстимульной импульсации нейронов в ряду сочетаний, как правило, не превышают 1,5-кратный уровень «фоновой» активности. Это означает, что в случае повышения в обучении возбудительного ответа регистрируется активация, которая в соответствии с упомянутыми ранее критериями не может считаться специфической. Этот факт, вместе с плавной динамикой ответа и наличием тормозных реакций у многих нейронов, свидетельствует, на наш взгляд, о том, что основная феноменология пластичности попадает в разряд изменения межсистемных отношений уже имевшихся до обучения элементов индивидуального опыта. Это вполне соответствует теоретическим положениям бихевиористов о том, что при выработке классических рефлексов и дифференцировок новой реакции, т.е. в терминах данной главы, нового акта поведения не формируется.
* * *
Изложенные ранее теоретические положения и экспериментальные данные позволяют в общих чертах описать основные моменты процесса научения с точки зрения системной психофизиологии.
В дефинитивном поведении циклически повторяющиеся соотношения организма со средой в виде адаптивных последовательностей поведенческих актов формируют отношения элементов опыта. За счет воздействия среды эти последовательности могут быть прерваны. В соответствии с имеющимся опытом вместо запланированного акта в таком случае реализуется ориентировочное поведение, сменяющееся следующим приспособительным актом. При многократном повторении такого воздействия происходит привыкание, т.е. сокращение перерыва в последовательности вплоть до его полного исчезновения. В случае когда «информационное» воздействие среды сопряжено с «подкрепляющим» в виде разрушающего воздействия либо появляющейся возможности удовлетворения «мотивированного» состояния, после ориентировочного совершается поведенческий акт, не входивший ранее в эту последовательность. При многократном повторении фиксируется новая последовательность смены поведенческих актов и обеспечивающие ее осуществление отношения элементов опыта.
Собственно научение, заключающееся в формировании нового элемента индивидуального опыта, начинается с возникновения проблемной ситуации, когда организм не может достигнуть желаемого результата за счет использования имеющихся в опыте функциональных систем. На нейрональном уровне этому соответствует длительное рассогласование метаболических потребностей клеток и синаптического притока (см. гл. 14). Это приводит к одновременной актуализации множества функциональных систем. Такая актуализация приводит в поисковом поведении к новым последовательностям поведенческих актов и изменениям набора прасистем, т.е. пробным соотношениям организма со средой. За счет имеющегося опыта поведения индивида в проблемных ситуациях этот набор не случаен, а подчинен опыту генерации успешных проб, накопленному индивидом. В случае неудачи состояние повышенной актуализации множества систем сохраняется и происходит генерация следующей пробы. В случае же успеха полученный результат снижает общий уровень актуализации систем, видимо, за счет частичного удовлетворения метаболических потребностей нейронов. После ряда успешных проб происходит исключение лишних конкурирующих альтернатив и окончательно складывается новая функциональная система поведенческого акта. Она имеет вид определенной организации актуализированных ранее сформированных систем и «добавки» группы специализированных нейронов, представляющих вновь сформированный элемент опыта в памяти организма. Эта «добавка» обеспечивает консолидацию кооперативного ансамбля, необходимого для достижения данного результата. Одновременно в силу включения нового акта в последовательность имевшихся поведенческих актов формируются отношения нового элемента опыта с уже имевшимися в памяти организма.
Дата добавления: 2016-06-13; просмотров: 813;