ЕСТЕСТВЕННЫЕ И ИСКУССТВЕННЫЕ ЖИЛИЩА ПЧЕЛ И ИХ СВОЙСТВА ПРИМЕНИТЕЛЬНО К ЗИМОВКЕ
Для жизни пчелиной семьи чрезвычайно большое значение имеет пчелиное жилище, которое должно соответствовать немалому количеству специфических требований, предъявляемых к нему семьей.
Что же касается пчеловода, то результаты его труда будут зависеть не только от знаний и умений, но и от того, настолько правильно и качественно спроектировано и изготовлено жилище для пчел — улей. В этой главе мы подробно рассмотрим все, что касается естественных и искусственных жилищ пчел.
Для однозначного понимания предмета наших исследований дадим определения для тех пчелиных жилищ, о которых будет идти речь дальше.
Дупло— замкнутая полость с отверстием в стволе живого дерева, образовавшаяся естественным образом вследствие разрушения древесины ядра из-за его гниения.
Борть— искусственное дупло в живом дереве, изготовленное человеком, использующееся для заселения пчелами при бортевом пчеловодстве.
Колода— простейший неразборный безрамочный улей, изготовленный из цельного толстого куска сухого дерева длиной до 1,5 м.
Улей — искусственное жилище пчел, изготовленное человеком из соотвехствующего материала (доска, камыш, пластмасса и др.), в котором пчелы строят свое гнездо на вынимающихся рамках.
Поскольку естественным первичным жилищем пчел было дупло, то и начнем с его рассмотрения.
► Дупло как жилище пчел
У многих пчеловодов давно стало аксиомой утверждение о том, что оптимальным жилищем для пчел является дупло. Упоминание об этом довольно часто встречается в литературе, однако мне неизвестен источник, где бы проводился системный анализ всех факторов, определяющих привлекательность дупла в качестве жилища для пчел. Давайте попытаемся провести такой анализ.
♦ Общие сведения о дупле
Как следует из данного выше определения, дупло представляет собой полость в стволе живого дерева, образовавшуюся в результате разрушения (гниения) древесины ядра.
|
Процесс гниения древесины — это биологическое разложение древесины дереворазрушающими грибами. Гниение древесины происходит с определенной скоростью от места заражения (механического повреждения, выпавшего сучка и т.д.) как вниз, так и вверх ствола и может достигать нескольких десятков сантиметров в год (Лесная энциклопедия, 1986). Полость дупла образуется в растущем живом дереве в результате ядровой гнили на последней ее стадии, когда происходит полное разрушение древесины ядра с постепенным формированием замкнутой полости. Однако после начала образования полости процесс гниения древесины ядра продолжается как в вертикальном направлении, что увеличивает высоту дупла, так и в горизонтальном, что увеличивает диаметр полости дупла.
В полностью сформировавшемся дупле гниение древесины ядра в вертикальном направлении прекращается при достижении полостью дупла корней. Гниение вверх может продолжаться и после заселения дупла пчелами, однако прополисование свода дупла и закрепление его восковыми сотами, видимо, предотвращает дальнейшее обрушение выгнивающей вверх древесины.
Что же касается гниения древесины ядра в горизонтальной плоскости, то этот процесс приостанавливается только после того, как в данном месте ядро дерева полностью разрушится и полость дупла дойдет до заболони. Древесина заболони устойчива к воздействию дереворазрушаю-щих грибов, вызывающих ядровую гниль, и поэтому не гниет. Часть пораженной, но не полностью разрушенной (ситовой) древесины ядра, примыкающей непосредственно к заболони, образует своеобразный переходной слой небольшой толщины. Ситовая древесина внутренней полости дупла на протяжении длительного времени сохраняет свою цельность, чему может способствовать ее умеренное прополисование после заселения дупла пчелами.
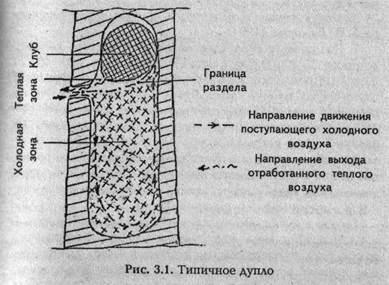
Если учесть все сказанное, то можно создать обобщенную модель типичного дупла (рис. 3.1).
Следует сказать со всей определенностью, что эта модель так же похожа на реальное дупло, как схематическое изображение человека на детских рисунках («ножки, ручки, огуречик, вот и вышел человечек») на фотографию конкретного человека. Это связано прежде всего с тем, что в природе двух одинаковых дупел, как и двух одинаковых людей, практически не бывает. Об этом более подробно мы будем вести речь дальше. Однако при помощи схематического изображения человека вполне возможно показать, как он ходит, бегает, прыгает и вообще — как он живет. Точно так же предлагаемая модель дупла вполне подходит для того, чтобы пояснить, как «живет» типичное дупло вместе с семьей пчел, разместившейся в его гостеприимной полости.
Ну а теперь давайте посмотрим, как выглядят реальные дупла в описании тех пчеловодов, которые их видели и изучали.
Начнем с описаний известного новгородского пчеловода М.В. Лупанова (1991).
1. Представьте себе толстую осину с дуплом, диаметр которого 35 см. Дупло длиной 4—5 м (объем 380—480 л) пчелы застроили сотами в 7 вертикальных рядов. Все ряды сотов сверху донизу прикреплены к стенкам дупла, но не сплошь, а с некоторыми промежутками: на протяжении 10—15 см соты прикреплены к дереву, а на протяжении 3—4 см — не прикреплены.
Пчелы строят соты в дупле обычно на холодный занос, что обеспечивает хорошую вентиляцию гнезда. Соты, построенные на теплый занос, можно встретить только тогда, когда в дупле имеется трещина (щель).
Летком в дупле обычно служит отверстие, образовавшееся в результате выпадения сгнившего сучка, или щель (трещина), искусно превращенная пчелами в леток. При заселении пчелами дупла и его обустройстве леток, как правило, находится ниже сотов, а потом при постепенной застройке дупла сотами он оказывается примерно посредине.
Дупло длиной 4—5 м пчелиная семья любой силы не может застроить сотами за 1—2 сезона, поэтому застройка происходит постепенно на протяжении нескольких лет.
2. Длина дупла в осине 5 м, диаметр — 30 см (объем 350 л), толщина стенок дупла — 15 см. Соты в нем располагались в 6 рядов. Длина сота, построенного за два сезона, была 150 см. Меда сотового в дупле было 55 кг.
3. Дупло в осине имело длину 5 м и диаметр 35 см (объем 480 л), соты были построены в 7 рядов на 4 м в длину. Соты построены на холодный занос. Леток находился посредине сотов с южной стороны дерева. Толщина
стенок дупла — 25 см. Дупло начиналось на высоте 2 м от земли. Сотового меда в нем было 10 ведер. Сверху сотов на 1 м мед был закристаллизован.
И вот еще что пишет М.В. Лупанов по поводу дупла: «Интересно, что в естественном жилище пчел — дупле всегда имеется благоприятный тепловой режим, так как там нет сквозняков. Воздух в дупло поступает только через леток и проникает во все уголки дупла без напора.
В зависимости от времени года и температуры пчелы сами регулируют движение воздуха. К зиме пчелы уменьшают леток, частично заделывая его прополисом, а к лету —' расширяют. Иногда в дупле, занятом пчелами, я обнаруживал трещину, но она всегда была тщательно заделана прополисом. Оставлено лишь небольшое отверстие для летка. И сот в таком дупле построен на теплый занос, т. е. устранена возможность сквозняка.
Интересно, что, сколько бы я ни снимал пчел с дупел, ни в одном дупле не видел подмора пчел. Дно дупла всегда было чистым.
Многолетние наблюдения убедили меня в том, что в природных условиях пчелы поселяются в просторных дуплах, в которых можно строить соты несколько лет и где имеется хороший обмен воздуха».

|
Ученый пчеловод Е.П. Петров (1983) из Башкирии в период с 1960 по 1968 г. вскрыл 12 дупел, по которым приводит обобщенную информацию (табл. 3.1).
Автор сообщает, что толщина стенок дупел колебалась от 3 до 20 см. Стенки изнутри имели слой трухлявой (ситовой) незапрополисованной древесины. Поэтому естественные жилища пчел в дуплах деревьев, как правило, хорошо сохраняют тепло и в них всегда сухо.
Как видно из таблицы, количество медовых запасов в гнездах пчел, зимующих в естественных дуплах, может быть самое различное — от 0,7 до 90 кг. Это зависит от силы семьи и срока заселения дупла. Сильные ранние рои собирают меда в два-три раза больше, чем поздние мелкие рои или изроившиеся материнские семьи. Иногда бывает, что в просторном естественном дупле мед постепенно накапливается годами за счет остатков неизрасходованных зимних кормов из запасов прошлых лет.
Е.П, Петров обращает внимание на своеобразие структуры пчелиного гнезда в условиях естественного существования семьи. Вся верхняя часть гнезда образует хранилище кормовых запасов, а ниже их располагается компактное гнездо с расплодом, сплетенное многочисленными перемычками между сотами. Такое строение создает единство, увеличивает внутреннюю компактность гнезда, расположенного в вертикальном пространстве, что способствует экономному расходованию корма и хорошему развитию семьи. При зимовке пчел в дупле большое значение имеет взаимное расположение запасов корма, расплода и летка. Между ними существует постоянное взаимодействие. В осенне-зимний период, когда развитие семьи прекращено полностью и расход корма пчелами ограничен до минимума, пчелиный клуб движется только вверх вслед за убывающими кормовыми запасами, но при этом «привязывается» к одному из летков дупла. По мере удаления от летка форма клуба меняется, и во второй половине зимовки он имеет чуть вытянутую форму, так чтобы край клуба всегда был по соседству с летком. Поскольку пчелы в зимнем клубе постоянно меняются местами, то каждая из них периодически попадает в зону притока свежего воздуха, идущего от летка. Следовательно, вопреки мнению некоторых современных исследователей, пчелы зимой все же нуждаются в постоянном притоке кислорода.
Относительно внутреннего устройства гнезда в дупле Е.П. Петров пишет: «Характерно, что в естественных дуплах... пчелы застраивают гнезда довольно беспорядочно. При вскрытии естественных жилищ пчел мы не видели ни одного гнезда, похожего на другое. В одних гнездах соты выстроены поперек летков, в других — наискось, одни имеют прямые соты, другие — волнистые, одни прикрепляют соты к стенке дупла язычком, другие — ребром. Причина такого разнообразия в постройках сотов — неодинаковая форма и место расположения летка: у одних семей летком служат отверстия от выпавших гнилых сучков, у других — трещина от молнии или мороза, в третьих — отверстие, выдолбленное дятлом, и т.д. Постройка сотов зависит также от силы семей. В целом же пчелы устраивают гнезда так, чтобы в них лучше сохранялось тепло.
...Верхняя и нижняя часть гнезда в дупле обычно заняты медовыми запасами.
...Наблюдения за семьями пчел в естественных дуплах показали, что иней, осевший в сильные морозы на стенках дупла, впоследствии исчезает, даже не оставляя сырости».
Сходную информацию относительно внутреннего устройства естественного гнезда в дуплах мне удалось обнаружить в «Русском пчеловодном листке» № 1 за 1903 год, в котором Д. Карчевский пишет: «Мне неоднократно приходилось видеть постройки диких пчел в дуплах деревьев. Из виденных мною естественных диких гнезд пчел нет двух гнезд, похожих друг на друга: одни из них строят соты поперек летка, другие наискось, одни имеют прямую, другие волнистую вощину (так тогда называли соты — В.К.),
одни строят одну пчелиную вощину, другие смешанную, одни прикрепляют соты к стенкам дупла язычком, другие — ребром».
Поразительное сходство этого описания с описанием Е. Петрова лишний раз подтверждает реальность существования таких явлений.
Относительно летков в естественных дуплах Д. Карчев-ский сообщает, что они имеют не только разные размеры и форму, но и размещаются или в самом верху дупла, или посредине, или гораздо ниже пчелиного гнезда.
Продолжаем знакомство с описаниями дупла. Информация от Василия Пуца (1997) с Волыни.
В марте месяце при валке леса в сосне было обнаружено дупло с пчелами. Гнездо пчел было высотой около метра. Внизу было семь сотов, а к верху сужалось до трех. Ориентировочный диаметр дупла внизу — 26 см, вверху — 12 см, объем — 120 л. Соты располагались выше летка. Ниже летка было 5 см пустоты, а затем шла трухлявая мягкая древесина. Толщина стенок живого дерева составляла всего 3,5 см. Медом в гнезде были заполнены около половины высоты сотов, весь он был запечатан. Перги не было. Соты внизу были черные, то есть в этом дупле пчелы жили давно.
Журнал «Паака» № 3 за 2005 год сообщает, что в 1947 г. в Киевской области было вскрыто дупло, диаметр которого составлял около 25 см, а высота — до 6 м. Меда в этом дупле было около б пудов. Соты были довольно чистыми светло-коричневого цвета.
Там же дается ссылка на Н.М. Витвицкого, который в свое время писал о дуплах, в которых находилось до 15— 20 пудов меда.
Журнал «Пчеловодство» № 9 за 1973 год сообщает о найденном в 1940 г. дупле в осиновом пне высотой 5,5 м. Высота сотов — 4,5 м, ширина — 28—30 см, масса пчел составляла 7 кг. Меда в дупле было 120кг. Все соты, как и стенки дупла, были чистыми, без следов поноса; подмора пчел на дне не было.
. Хочу поделиться также и своими наблюдениями. В Богодуховском районе Харьковской области в дикой груше высотой около 20 м и диаметром у основания 65 см было обнаружено дупло (рис. 3.2).
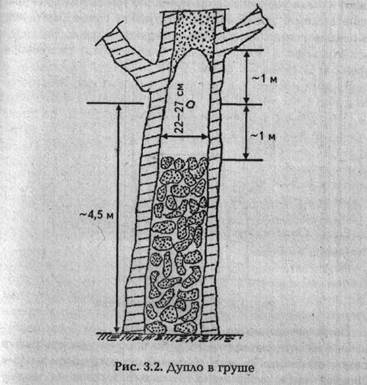
Внутренняя часть ствола до высоты около 6 м выгнила. Внизу толщина оставшейся живой древесины — 10—12 см,в районе дупла — 5—12 см. Пустота, в которой жили пчелы, имела высоту 2 м. Леток диаметром 6 см был расположен на высоте около 4,5 м от земли в направлении на юг. Леток делит полость дупла на две равные части по 1 м. Объем внутренней полости дупла около 100 л. Ниже внутренней пустоты выгнившая часть ствола до самой земли была плотно заполнена кусками пористой трухлявой древесины (ситовины). Ситовина была и выше верхнего свода дупла, но там она была скреплена с живой древесиной и разрушена только частично. В районе гнезда внутренняя поверхность дупла незапро-полисованная и почти гладкая. Слой плотной ситовины в этой части дупла не более 5—10 мм. Непосредственно у летка дупло имело вид, представленный на рис. 3.3.
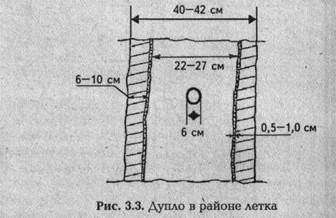
На момент осмотра дупла все соты в нем были разрушены и на внутренних стенках дупла просматривались только следы от сотов. По этим следам было установлено, что в средней части дупла было построено 5 сотов на теплый занос. Вверху количество сотов уменьшалось до трех. Создавалось впечатление, что в оголовке соты прикреплялись не прямо к своду дупла (ситовине), а держались на носковых перемычках длиной 8—10 см (по диаметру верх- ней части), прикрепленных к стенкам дупла. Трухлявая си-товина вверху дупла сохраняла целостность, но прочность ее была весьма низкой.
Леток правильной формы диаметром 6 см образовался в результате выживания сучка. Толщина живой древесины (заболони) в районе летка — около 5 см. В нижней части дупла было еще несколько отверстий и трещин, но все они были изнутри дупла закрыты кусками пористой трухлявой древесины.
Продолжая сбор информации, остановимся на общих описаниях дупла, опубликованных в разных источниках.
Е.К. Еськов (1992) пишет: «Естественное пчелиное гнездо представляет собой довольно беспорядочную систему лабиринтов из сотов, которые различаются по форме, размерам, прикреплению к гнездовой полости и друг к другу... Беспорядочное, размещение сотов в гнезде медоносных пчел объясняется тем, что на начальных этапах строительства при освоении новой полости для жилья они не учитывают перспективы гнездовых построек.
При возможности выбора пчелы вселяются преимущественно в полости, ранее занимаемые другими семьями. Предпочитаемый объем полости 30—80 л. Предпочитают селиться на опушках леса и других освещенных солнцем местах, а также вблизи водоемов. В большинстве случаев пчелы выбирают дупла на высоте до 3 м с летком, обращенным на юго-запад».
Ю.К. Барбарович (1993) отмечает: «Все ряды сотов сверху донизу прикреплены к стенкам дупла, но не сплошь, а с некоторыми промежутками: на протяжении 10—15 см соты прикреплены к дереву, а на протяжении 3—4 см — не прикреплены и образуют отверстия для прохода пчел. Обычно в дупле бывает 7—8 сотов. Пчелы строят соты в дупле обычно на холодный занос, но можно встретить и теплый занос, и соты под углом.
Ситовина внутренней полости дупла, как губка, вбирает большое количество влаги, так как ее гигроскопичность в 3—5 раз выше, чем у живой древесины».
С. Клименков в журнале «Пчеловодство» № 9, 1973 на с. 23 сообщает, что в 1940 г. он нашел гнездо пчел в осиновом дупле высотой 5,5 м. Леток диаметром 40 мм был с южной стороны в 4 м от земли. Соты размещались на холодный занос, их высота превышала 4,5 м, а ширина составляла 28—30 см. Сотов с медом было вырезано 120 кг. Масса пчел 7 кг. Стенки дупла были чистыми, подмора на дне не было.
И.А. Шабаршов (1996) пишет: «В живом дереве, в силу протекающих естественных процессов, летом пчелам не так жарко, а зимой не так холодно, как в засохшем мертвом дереве. Сами пчелы инстинктивно выбирают себе дупла в живых деревьях. Кстати, до сих пор остается загадкой, чем они руководствуются, бракуя дупла в сухих деревьях.
В живом дереве, в котором и зимой не приостанавливаются процессы обмена, ни холод, ни сырость не могут вредить пчелам».
По утверждению Н.М. Витвицкого, зимующие в дуплах пчелы не испытывают недостатка в свежем воздухе. Они сами научились регулировать его доступ через летки. В сильной, богатой медом семье пчелы на зиму оставляют обычно большой просторный леток. Семья, не очень сильная осенью, его уменьшает, но делает таким, что через него проходит в жилище столько воздуха, сколько ей нужно. Такую закономерность Витвицкий наблюдал почти в каждом дупле и по размеру зимнего летка мог безошибочно судить о силе семьи.
Т. Цесельский (2001) отмечает: «Наиболее подходящими помещениями являются для пчел дупла живых деревьев, так как циркулирующие в дереве соки способствуют поддержанию умеренной температуры во время летней жары, в зимнюю же пору живые слои древесины препят-
ствуют доступу сильного мороза к внутренним слоям дерева. Поэтому, как показывает опыт, пчелы с особой охотой устраивают свое жилище в таких дуплах, и в них существование семьи поддерживается лучше всего. Дупла мертвых деревьев менее удобны, почему и случается довольно часто, что пчелы, долгие годы благополучно живущие в дупле живого дерева, погибают вскоре после того, как дерево засохнет».
М.И. Кошкин (2003) обобщил информацию о 130 дуплах в Уссурийском крае и на Чаткале. Он пишет о том, что пчелы отдают предпочтение дуплам, находящимся в деревьях, расположенных на склонах южной четверти. Какой-либо твердой закономерности в расположении сотов в дупле относительно сторон света им не установлено. Гипотеза о геомагнитной ориентации сотов в дуплах не нашла подтверждения. Не обнаружено также никакой закономерности в расположении сотов относительно теплого или холодного заноса.
Сообщения о многометровых сотах в дуплах и многопудовых запасах меда — «сказки чистейшей воды». В природе автор не встречал семьи, способные собирать и запасать такое огромное количество меда. «Живя в дупле, пчелы летом собирают мед, который к весне полностью расходуют, так что многолетних накоплений тут быть не может».
В.В. Родионов и И.А. Шабаршов (1968), освещая данный вопрос, пишут: «Пчелиная семья поселяется обычно в верхней части дупла. К его потолку пчелы прикрепляют соты. Какой бы величины соты ни были, под ними почти всегда остается пустое, незанятое пространство. Нижнюю часть стенок и дно дупла пчелы никогда не прополисуют. Леток у них всегда открыт, только при теплой и умеренной зиме он бывает более широким, а в суровую и продолжительную — небольшим.
Углекислый газ, насыщенный водяными парами, который выделяется в результате жизнедеятельности семьи, в силу своих физических свойств опускается вниз дупла. Здесь он, как губкой, поглощается и удерживается трухлявой пористой древесиной. Ее поглотительные свойства тем выше, чем больше дупло. Удаление влаги в дупле — процесс не только механический. Он, несомненно, связан сжизнедеятельностью микрофлоры, разрушающей гниющее дерево. Видимо, на пчел благоприятно влияет само дерево, в котором, как известно, жизненные процессы не прекращаются и зимой. Примечательно, что пчелы строят гнезда только в дуплах живых деревьев.
То количество кислорода, которое требуется пчелам зимой, поступает к ним через леток и через него же выходит из дупла часть паров и углекислого газа. Леток прекрасно выполняет роль вентилятора».
Итак, пока этой информации будет вполне достаточно для того, чтобы представить себе, какое же оно есть на самом деле — это такое простое, но вместе с тем и загадочное дупло. Теперь попытаемся проанализировать и систематизировать всю эту информацию для того, чтобы создать портрет наиболее часто встречающегося (типичного) дупла.
— Дерево.Первичное заселение пчелы проводят только в дупла живых деревьев.Если в процессе жизни
семьи дерево засыхает, то пчелы или слетают с него, или погибают. Есть множество подтверждений этому факту. Одно из них приводит В. Ливенцев, который описывает факт гибели зимующей пчелиной семьи после того, как засохла осина, в которой размещалось дупло с пчелами.
— Порода дерева.Подавляющее большинство пчелиных семей селятся в дупла, находящиеся в деревьях
лиственных пород.Это связано с тем, что деревьяэтой породы' в гораздо большей степени подвержены такому пороку, как ядровая гниль, в результате которой образуются дупла. В деревьях хвойных пород дупла встречаются очень редко. В 18 случаях, которые наблюдал В.Д. Крутоголов (2002), пчелы отдавали предпочтение дуплам в дубах и липах.
— Леток.Чаще всего располагается в районе средней части дупла,а под летком, как правило, есть свободное пространство, своеобразный воздухообменник, иногда значительного объема. Форма летка может быть самой разнообразной в зависимости от происхождения летка, но чаще всего он имеет круглую форму (выгнивший сучок, вход, проделанный мышью, дятлом и т.д.). Но даже если вход в дупло представляет щель в дереве, пчелы, как правило, за делывают эту щель воском и прополисом, оставляя только круглое отверстие необходимого диаметра.
— Внутреннее покрытие полости.Внутри дупла между свободным пространством и живой древесиной
заболони имеется тонкий слой пористой трухлявой древесины (ситовины).
— Процесс разрушения древесины.После образования дупла и заселения его пчелами процесс разрушения (гниения) древесины ядра может продолжаться дальше.Однако в горизонтальной плоскости этот процесс останавливается, когда разрушение достигнет живой древесины заболони. В вертикальной плоскости процесс гниения древесины может продолжаться еще длительное время.
— Прополисование внутренних стенок дупла.По этому вопросу на сегодня нет единого мнения. Одни пчеловоды утверждают, что пчелы обязательно прополисуют дупло, другие сообщают, что дупло имеет незапрополисованные стенки. Что касается меня, то я склонен разделить последнее мнение. Тот микроскопический слой прополиса, который удалось обнаружить в дуп-ле в груше, я бы не назвал прополисованием дупла для его герметизации. Скорее всего, этот тончайший слой прополиса выполнял санитарную функцию для стерилизациивоздуха в дупле и защиты пчел от воз-будителей болезней.
Вполне допустимо, что в определенных районах (например, на севере) или в определенных породах деревьев пчелы ; все же прополисуют дупло. Но в таком случае они пропо- ; лисуют только верхнюю часть дупла, где располагается семья. Нижнюю часть дупла пчелы никогда не прополисуют.
В подтверждение этой версии приведем информацию упомянутого выше В.Д. Крутоголова: «Заметил, что пчелы прополисуют дупло только там, где расположено гнездо... В других местах они лишь очищают дупло от частиц гнилой древесины, которые могут осыпаться».
Е.П. Петров (1983), исследовавший в Башкирии 12 дупел в живых деревьях, сообщает: «Стенки изнутри (дупел) имеют слой трухлявой (ситовой) незапрополисованной древесины. Поэтому естественные жилища пчел в дуплах деревьев, как правило, хорошо сохраняют тепло и в них всегда сухо».
Г.Ф. Таранов в журнале «Пчеловодство» № 4 за 1982 год пишет: «Если верх дупла пчелы прополисуют, то боковые стенки бывают открыты, и через них проникает воздух и водяные пары».
Разночтения в вопросе о прополисовании дупла могут возникать также и по той причине, что разные породы пчел, и даже разные семьи одной породы, могут по-разному про-полисовать дупла. Опытные пчеловоды хорошо знают, что есть семьи, которые очень сильно прополисуют улей, а есть и такие, которые делают это средне и даже слабо.
— Пространственная ориентация сотов.Наблюдения показывают, что она не отличается строгой упорядоченностью.Утверждение отдельных ученых о том, что при свободной постройке сотов пчелы привязывают их направление к силовым линиям магнитного поля Земли, не находит подтверждения в жизни. Многообразие формы естественных жилищ, к которой пчелы и привязываются при строительстве сотов, значит для них гораздо больше, чем магнитная ориентация.
Мне самому приходилось многократно видеть свободно построенные соты. Какой-либо закономерности в их ориентации я не установил. А для некоторых построек понятие «ориентация» вообще неприемлемо: волнообразно извивающиеся соты скорее похожи на меха гармони, чем на упорядоченную постройку. Поэтому я полностью согласен с приведенными выше утверждениями Д. Карчевского и Е. Петрова о том, что «...нет двух гнезд, похожих друг на друга». Следовательно, есть все основания полагать, что в дупле соты могут быть построены как на холодный, так и на теплый занос, а также под углом к летку.Единственная константа, которая четко просматривается во всех этих постройках, это расстояние между смежными сотами — ширина улочки. У свежеотстроенных сотов она равна 12—13 мм, а по мере старения сотов за счет их достройки ширина улочки может значительно уменьшаться (до 5—6 мм). Но такие гнезда пчелы, как правило, покидают.
Характерной особенностью сотов в дупле является их неразрывностьв вертикальной плоскости, чего не бывает в наших ульях с рамками.
Общая площадь сотов (сумма площади двух сторон) может превышать 3 м2 (Еськов Е.К., 1991).
— Крепление сотов в дупле.Основное крепление сотов осуществляется прежде всего к стенкам.Что же касается прикрепления к потолку, то иногда в дуплах,где процесс разрушения ядра продолжается, пчелы; соты с потолком не соединяют. В этом случае верх сотов крепится к своеобразным восковым перемычкам, которые двумя своими концами присоединяются: к стенкам вверху дупла.
Соты прикрепляются к стенкам дупла не сплошь, а с некоторыми промежутками: на протяжении 10—15 см соты присоединяются к стенкам, а на протяжении 3—4 см пчелы оставляют пустоты. Через эти щели пчелы могут свободно переходить в любую улочку гнезда.
— Вдупле не может быть никаких сквозняков,так как свежий воздух поступает, а использованный удаляется из замкнутой полости дупла через один леток.
— После зимовки на дне дупла не бывает подмора.Эту особенность пчелиного гнезда в дупле живого дерева отмечают многие наблюдатели.
— Замкнутая полость дупла в живом дереве является отличным экраном для внешних электрических полей естественного и искусственного происхождения. По этой причине в гнезде пчел внутри дупла нет негативного воздействия электрополей.
— Стенки дупла живого дерева, в котором постоянно находится вода с питательными элементами, являются своеобразной ловушкой для теплового излучения пчелиной семьи. Это связано с тем, что слой воды толщиной всего в несколько сантиметров является непрозрачным для теплового (инфракрасного) излучения. Следовательно, вода (пасока) живого дерева летом защищает гнездо от внешнего перегрева солнечными лучами, а зимой способствует отражению и сохранению внутреннего тепла зимнего клуба.
По всей видимости, это обстоятельство и является одной из причин того, что пчелы отдают предпочтение новому жилищу в дупле живого дерева, а не в сухостое.
Все сказанное выше относится к типичному дуплуоптимального объема и формы. Дупла-малютки менее этого объема, а также дупла другой формы (нетипичные дупла) не могут обеспечить необходимых условий для нормальной жизнедеятельности пчелиной семьи, и поэтому к ним не относится тот анализ, который мы будем проводить дальше. Заметим более определенно — нетипичные дупла никогда не могут быть оптимальным жилищем для пчел. А если пчелы в них все же поселяются, то это — вынужденный вариант для пчел, которые просто не смогли найти себе более подходящего жилища.
Исходя из этого, предлагаю разделить дупла на типичные(о которых мы и ведем речь) и нетипичные.Кстати, полярные отношения некоторых пчеловодов к дуплу как жилищу пчел, мне кажется, как раз и объясняются тем, что они под термином «дупло» понимают совсем разные физические жилища. И поэтому можно вести бесконечные споры (что мы и видим в пчеловодной литературе как минимум последней сотни лет), не понимать друг друга и оставаться при своем мнении. А дискуссия-то эта — о совсем разных вещах. С таким же успехом можно вести спор о свойствах дерева, не обозначив предварительно, о каком дереве ведется речь. Ведь спор об эбеновом «дереве» или о бальзовом «дереве» беспредметен без их предварительного обозначения — одно дерево (эбеновое) тонет в воде, а другое (бальзовое) легче пробки.
Возвращаясь к дуплу, скажу, что именно таким я и представляю себе на сегодняшний день типичное дупло — жилище пчел в живом дереве.
♦ Естественный закон Ф.А. Соколова
Однако представленная информация о дупле была бы неполной, если бы я не представил еще одну точку зрения. Принадлежит она Федору Алексеевичу Соколову, выдающемуся русскому пчеловоду, который еще на заре XX в. в журнале «Русский пчеловод» представил ее в виде «Естественного закона о жизни дикого роя». Это были результаты его тридцатилетних исследований жизни пчел в дуплах живых деревьев.
Цитирование приводится по книге Б.В. Крюкова и К.И. Носовой «Пчелы выбирают дупло» (1995 г.).
1. Всякое дерево, достигая известного возраста и старости, теряет свою сердцевину... которая усыхает,
разрушается и сгнивает, а после этого внутри дерева образуется в большинстве случаев пустое пространство, имеющее направление сверху ствола дерева вниз... называемое нами дуплом.
2. Всякое дерево, имеющее дупло, но здоровую древесину (наслоение, окружающее сердцевину), продолжает не только жить, но и плодоносить.
3. Пустота или дупло... всегда имеет одинаковый характерный вид: оно начинается на известной высоте выгнившей сердцевины дерева незначительною пустотою,вроде 1—2 вершка (1 верш. = 4,5 см. — В.К.) в диаметре, направляется вниз к корню дерева и образует там большую ширину и глубину пространства, чем толще старое дерево, а при этом дупло постепенно расширяется внутри дерева книзу, сообразно сгнившей и выпавшей сердцевины, т. е. образует пустое, конусообразное пространство, узкое вверху и широкое внизу.
4. Всякий рой, входя в дупло дерева, направляется непременно вверх, то есть в голову дупла, там свивается, очищает потолок от сора и там... начинает свои восковые постройки... матка приступает к кладке яичек в середине строящихся роем пластов вощины...(так в те времена называли соты. — В.К.).
5. По мере построек вощин вниз матка тоже опускается за роем вниз дупла для кладки яиц в вощинах, но с наступлением первых же холодных ночей отступает опять вверх, а рой, по мере выхода молодой пчелы, тоже отступает постепенно вверх дупла дерева.
6. По мере выхода молодой пчелы в голове дупла дерева, рой заливает освободившиеся ячейки вощин медом и печатает их, чем преграждает матке новую кладку яиц в эти же ячейки. Таким образом, в голове дупла дерева скапливается постепенно запас печатного меда, необходимого рою для зимнего довольствия.
7. Иногда сильный рой выстраивает восковые постройки далеко ниже входного отверстия в дупло дерева и тоже заливает их медом, но никогда не остается здесь, под медом, на зиму, т.е. ниже отверстия или летка, а избирает для этого в дупле дерева выше летка такое место, которое в состоянии закупорить своею массою поперек, чтобы не допустить мимо себя несоответствующей температуры воздуха, который может охладить мед, сделать его... негодным для употребления.
8. Обеспечив снизу своею массой правильное хранение кормового меда, рой заботится также и о том, чтобы из головы дерева не уходил теплый воздух, выделяемый клубом роя, а потому еще летом плотно и наглухо заклеивает голову и стенки дупла дерева прополисом, вплоть до окончания места, избранного роем в дупле дерева для зимнего своего пребывания.
9. Рой не заклеивает входного отверстия в дупло дерева, или летка, в продолжение всего лета, но после наступления холодных ночей обязательно заклеивает леток и тем больше, чем более увеличивается дневной и ночной холод к осени. Затем, к тому времени, когда температура воздуха внешнего держится еще около +8 °С по Реомюру (10 X), рой уже прекращает ход в поле и заклеивает леток прополисом настолько, насколько рою необходим лишь малый приток в дупло дерева холодного, но чистого воздуха для замены теплого, но испорченного дыханием пчел воздуха.
10. Относительно большей или меньшей степени заклейки летка на зиму всякий рой соображается, прежде всего, с собственною силою, а потом с обширностью и глубиною находящегося под ним пустого пространства в дупле дерева, и чем короче и уже пустое пространство в дупле дерева под роем, тем меньше рой заклеивает леток.
11. Не имея никакой возможности регулировать леток зимой, рой, заклеивши его с осени, до известной степени регулирует зимой внутри дупла самый воздух над собою посредством сокращения или расширения своего клуба и под собой посредством движения крыльев, то, поднимая температуру, то, опуская ее, когда это нужно.
12. Всякий дикий рой защищен от чрезмерного холода со стороны стенок дупла дерева тем, что всякое живое дерево, в силу своей организации не замерзает зимой ни при каких морозах, а только теряет теплоту до известного предела, колеблющегося между + 1—2 °С по Реомюру (1,25—2,5 °С).
(Примечание.Это утверждение Ф.А. Соколова требует дальнейшего уточнения и проверки в натурном эксперименте. — В.К.)
13. Холодный и тяжелый воздух, входя в дупло дерева через нижнюю половину летка, направляется не вверх, прямо к клубу зимующего роя, а течет по передней стенке дупла дерева вниз, на дно дупла, здесь смешивается, не производя ни малейшей испарины, постепенно согревается, становится от этого более легким и потому постепенно поднимается вверх, к рою и снабжает его чистым воздухом для дыхания. Из этого ясно видно, что, чем глубже дупло дерева под роем, тем лучше, здоровее и спокойней зимует в нем всякий дикий рой.
14. Хотя всякий дикий рой и подвергается с осени до весны оцепенению в силу своей физиологической организации, но не полному оцепенению, а потому живет и до некоторой степени бодрствует, питаясь медом, как пищей и питьем, а, равно регулируя воздух как над собой, в голове дупла, так и под собой, до дна дупла дерева, теми способами, какие указаны выше.
15. Ведя в продолжение осени и зимы такую совершенно естественную и вполне нормальную жизнь, дикий рой благополучно доживает до весны, выходит весной в целости и сохранности и теряет лишь несколько старых и больных пчелок, которые упадут на дно дупла дерева.
А вот теперь мы располагаем достаточным количеством информации для того, чтобы определить, является ли дупло в живом дереве оптимальным (наилучшим) жилищем для пчел или нет.
♦ Анализ свойств дупла как жилища для пчел
Чтобы проанализировать соответствие дупла оптимальному жилищу для пчел, сформулируем основные требования к такому жилищу:
— Обеспечение необходимого для жизнедеятельности семьи замкнутого физического объема с отверстием
(летком).
— Защита от неблагоприятных факторов внешней среды и прежде всего от дождя, снега, ветра.
— Надежная теплоизоляция, т. е. защита от перегрева и охлаждения.
— Обеспечение семьи свежим воздухом, точнее — кислородом.
— Удаление из гнезда продуктов жизнедеятельности пчел и прежде всего влаги и углекислого газа.
О второстепенных требованиях, таких как механическая прочность, защита от хищников и вредителей и др. здесь говорить не будем.
Из анализа представленной выше информации следует, что первые четыре требования в дупле, как правило, выполняются. А вот как реально происходит удаление продуктов жизнедеятельности семьи и особенно из зимнего клуба — не совсем ясно.
В некоторых источниках есть, правда, объяснения, что влага из дупла зимой удаляется за счет диффузии: постут пающий через леток в дупло морозный сухой воздух поглощает влагу, выделяемую клубом. Но ведь и в улье можно создать механическое подобие дупла, поставив под зимовальный корпус пустой магазин или корпус, загерметизировав потолок, закрыв нижний леток и открыв верхний. Условия для диффузии в этом случае будут, как в дупле, однако, хотя такой вариант зимовки является приемлемым, все же назвать его оптимальным нельзя, прежде всего по причине высокой влажности в гнезде.
Значит, влага из дупла, помимо диффузии, удаляется еще каким-то другим способом? Вентиляцией? Но ведь сквозняков в дупле нет, так как там всего один леток. Следовательно, существует еще один какой-то механизм удаления влаги. Еще начиная с Н.М. Витвицкого и до некоторых авторов наших дней высказывается предположение, что само живое дерево каким-то образом участвует в удалении излишков влаги из дупла, однако описания действия этого механизма нет. С.А. Поправко (1985) по этому поводу пишет: «Живое дерево облегчает пчелам существование в зимнее время, активно адсорбируя избыточную влагу через стенки дупла». Б.В. Крюков, К.И. Носова, Е.П. Петров эту версию вообще отрицают.
И что уж совершенно точно, так это то, что в современной пчеловодческой литературе по этому вопросу нет устоявшегося мнения, а существует явный информационный голод. Давайте попробуем разобраться в этом непростом, но интересном вопросе. При этом раскрытие и описание механизма удаления влаги будем делать для самого трудного для пчел периода — зимовки.
Но для дальнейших рассуждений в этом направлении надо вспомнить, хотя бы в общих чертах, как устроено живое дерево и какие функции выполняет оно для обеспечения своей жизнедеятельности.
♦ Взаимодействие основных элементов живого дерева
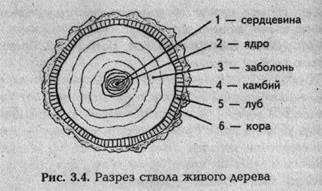
Основные элементы живого дерева — корневая система, ствол и крона. В самом центре ствола расположено цилиндрическое образование малого радиуса — сердцевина, которую опоясывает ядро (спелая древесина), за которой следует молодая древесина — заболонь. Внешние контуры ствола окаймляет кора. Между заболонью и корой находятся камбий (ближе к заболони) и луб (ближе к коре) (рис. 3.4.)
Рост ствола в толщину начинается с камбия — особой образовательной ткани, которая дает начало проводящим тканям — древесине и лубу. Камбий с определенной последовательностью откладывает внутрь новые слои древесины (годичные кольца), а кнаружи — луб. Наивысшая активность камбия наблюдается весной. Совокупность тканей между сердцевиной и камбием принято называть древесиной. Живая
древесина для обеспечения своей жизнедеятельности выполняет следующие функции:
— механическую;
— запасающую;
— проводящую.
Механическая функцияживой древесины обеспечивает механическую опору для кроны и самого ствола.
Запасающая функцияобеспечивает запас необходимых питательных веществ и влаги для жизнедеятельности дерева.
Проводящая функцияживой древесины состоит в ее свойстве перемещать (проводить) влагу и питательные вещества по всему своему объему.
Какие же элементы дерева обеспечивают реализацию этих функций?
Заболонь, являясь физиологически активной частью древесины, обеспечивает перемещение влаги и питательных веществ от корней вверх по стволу. В заболони содержатся не только одревесневшие (мертвые) клетки, выполняющие механическую функцию, но и живые (паренхимные) клетки, которые обеспечивают проводящую функцию и в которых откладываются запасаемые питательные вещества. Заболонь обладает высокой проницаемостью для влаги в любом виде — как для пара, так и для воды.
Ядро состоит из одревесневших (мертвых) клеток, которые образуются в процессе роста и жизнедеятельности дерева из живых клеток, ранее принадлежавших заболони. В процессе их отмирания происходит отложение ядровых веществ и закупорка водопроводящих элементов. В соответствии с этим ядро отличается от заболони более низкой влажностью, повышенным содержанием экстрактивных веществ и резко сниженной водо- и газопроницаемостью.
Древесина ядра более плотная, чем заболони, и более прочная, поэтому в дереве она выполняет основную механическую функцию. В растущем дереве ядро легче поражается дереворазрушающими грибами и гнилями, чем заболонь.
Ядро и заболонь в перпендикулярном (горизонтальном) направлении пронизывают сердцевинные лучи, которые обеспечивают перемещение влаги, питательных веществ от центра дерева к его периферии. Есть утверждения, что по сердцевинным лучам происходит также и воздухообмен в горизонтальном направлении (ж. Паака № 3, 2005).
Как известно, дупло в деревьях лиственных пород образуется в результате ядровой гнили на последней ее стадии, когда происходит полное разрушение древесины ядра. Гниение древесины происходит с определенной скоростью от места заражения (где впоследствии образуется леток) как вниз, так и вверх ствола и может достигать нескольких десятков сантиметров в год (Лесная энциклопедия, 1986). В соответствии с расположением дупла в дереве и свойствами древесины, дупло как жилище пчел имеет практически непроницаемый для влаги свод в виде нижней части разрушенного ядра, а по бокам дупло опоясывает заболонь, через которую легко проникает влага. По наружному контуру пустоту дупла окаймляет тонкий слой пористой трухлявой древесины (ситы), которая хорошо впитывает влагу. По опоясывающей дупло живой заболони происходит постоянный ток влаги от корней вверх по стволу. Проводящими элементами в заболони являются в основном так называемые трахеи (сосуды), образованные живыми клетками древесины, которые в своей оболочке имеют поры для улучшения механизма водопроведения. Соединяясь между собой, тысячи клеток образуют вытянутую вдоль по стволу сквозную трубку. Множество трубок соединяются своими боковыми частями в проводящие пучки, которые тянутся от корня по стволу вплоть до кончиков ветвей и листьев.
Как следует из сказанного выше, основой проводящих элементов древесины является клетка. Она состоит из оболочки и внутренней части — протопласта, состоящего в свою очередь из ядра и цитоплазмы. Клеточная оболочка (стенка) имеет полую структуру из нескольких слоев. Взаимодействие между клетками осуществляется через пустоты в оболочке. Пустоты бывают двух видов: мелкие — поры и более крупные отверстия в стенках клеток — перфорации. Так как живые древесные клетки имеют довольно большую протяженность — до 80 мм и даже больше, то в каждой клетке может быть до 300 пор, через которые осуществляется взаимодействие между самими клетками и взаимодействие с межклеточной средой. Перфорации находятся в основном в нижней и верхней частях клеток и обеспечивают проводящие функции клеток.
Неотъемлемым свойством любых живых клеток является их способность запасать воду и питательные вещества. Всякий дефицит воды в клетке ведет к созданию внутриклеточного разрежения и «засасыванию» воды и питательных веществ из межклеточного пространства. При полном насыщении клетки избыточное давление внутри нее может достигать 15—20-ти (Лесная энциклопедия, 1986).
Ранней весной с началом потепления при прогреве ствола возобновляется интенсивное сокодвижение, в первую очередь за счет запасов свободной влаги, хранящейся зимой в межклеточных пространствах. По мере прогрева почвы и корнеобитаемого слоя возобновляется поступление влаги от корней, включается механизм активного поглощения воды древесным организмом. Движущей силой активного поглощения является так называемое корневое давление. Механизм активного поглощения действует только ранней весной. Второй механизм, вызывающий пассивное поглощение влаги, определяется тем, что вода, заключенная в тонкие капилляры (сосуды, трахеи) проводящей системы и обладающая большой силой молекулярного сцепления, самопроизвольно движется в дереве по градиенту водного потенциала — от высокого в почве к низкому в атмосфере. Механизм пассивного поглощения влаги действует практически на протяжении всего вегетационного периода.
А куда же расходуется вода в дереве? Прежде всего, на поддержание постоянства внутренней среды дерева, т. е. на насыщение клеток и межклеточного пространства водой, а также на испарение (транспирацию). Основной орган транспирации — лист. Количество воды, испаряемое 1 г листьев в час, может достигать 1—1,5 г (Лесная энциклопедия, 1986). Если в зрелом дереве листовая масса составляет несколько десятков килограммов, то водопотребле-ние такого дерева летом может доходить до 200—300 кг воды в сутки. Ствол такого дерева летом представляет буквально «водопроводную систему». При сброшенной листве транспирация идет через покровные ткани (так называемая кутикулярная транспирация), интенсивность которой в 10—20 раз ниже листьевой. Потеря воды при транспирации, обусловленной сосущей силой атмосферы, создает дефицит давления в сосудах, которое передается через гидростатическую проводящую систему вниз до самих корней. Однако если на пути передаваемого вниз дефицита давления в сосудах появится посторонний источник влаги, то эта влага будет засасываться в проводящую систему дерева из этого источника еще до поступления влаги из корней.
О других особенностях функционирования живого дерева в период осень — зима — весна более подробно рассказано в приложении 1.
♦ Версия влагообменных процессов в дупле с пчелиным гнездом
А теперь вернемся к дуплу с находящимся в нем пчелиным гнездом.
Известно, что в процессе своей жизнедеятельности пчелы потребляют мед, в результате чего выделяют так называемую метаболическую воду. Потребляя 1 кг меда, пчелы выделяют около 700 г воды. Избыток воды, выделяемый пчелиной семьёй, накапливается в дупле. Часть этой влаги удаляется через леток, а часть — поглощается пограничным слоем трухлявой пористой древесины дупла. Поскольку пчелы выделяют влагу постоянно, то подпитка пограничного слоя влагой будет тоже идти постоянно. Следовательно, этот слой с накопленной в нем влагой можно рассматривать как посторонний источник влаги для дерева.
Поскольку к пограничному слою примыкает живая древесина заболони с открытыми концами (перфорациями), а также с боковыми порами сосудов и трахей, то через них влага из пограничного слоя (постороннего источника) будет засасываться в проводящую систему дерева и в нем использоваться по назначению (рис. 3.5).
Образование влаги в виде воды из влажного теплого воздуха клуба происходит в ситовине, примыкающей непосредственно к гнезду, за счет разности температур At°.
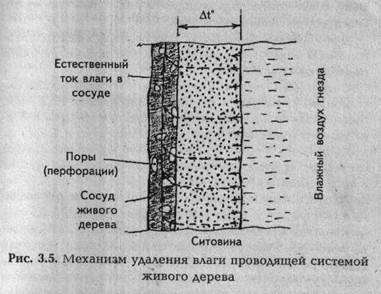
Имеется в виду то, что температура выходящего из клуба воздуха всегда положительна (в районе 10 °С), а температура заболони зимой будет ниже этой температуры. В таком случае влага, которую самопроизвольно всасывает сита по причине своей пористой структуры, по мере приближения к заболони в какой-то момент переходит температуру точки росы и начинает конденсироваться внутри ситовины, превращаясь в воду. Но все же основное количество влаги из отработанного теплого воздуха клуба образуется при его соприкосновении с холодной массой воздуха в нижней части дупла ниже гнезда. Влага, выпадающая при этом в виде росы или инея, будет впитываться ситовиной (роса) или оседать в нижней части дупла в виде наледи (иней). В первом случае влага будет удаляться проводящей системой дерева, кода она работает, а при очень низких температурах (экзогенный покой) эта влага будет также превращаться в лед и там (внизу) сохраняться до первых потеплений.
Для удобства дальнейших рассуждений назовем предложенный механизм удаления влаги так: «удаление влаги проводящей системой».
С действием механизма удаления влаги проводящей системой мы встречаемся довольно часто в быту. Например, таким образом идет потребление воды срезанными цветами или черенками винограда, которые мы ставим в воду на проращивание. Ну а уж полная аналогия будет при посадке срезанного черенка винограда, черной смородины, малины и др. в землю. В этом случае до образования корней (а это довольно продолжительный процесс) черенки будут потреблять (удалять) воду прямо из влажной земли за счет действия механизма удаления влаги проводящей системой.
Подобный механизм поглощения влаги существует и в неорганическом мире. Так, например, такие осушители, как цеолит, силикагель, поглощают влагу непосредственно из воздуха, даже минуя превращение влаги в жидкую фазу воды.
Подводя итог сказанному выше, можно сделать вывод о том, что живое дерево на протяжении вегетационного периода является постоянным потребителем и регулятором влаги в дупле, заселенном пчелами.
А как же будут обстоять дела зимой, когда деревья лиственных пород, в дуплах которых в основном и селятся пчелы, сбрасывают листву и переходят в состояние покоя? Оказывается, в живой древесине даже в состоянии глубокого зимнего покоя обменные процессы полностью не прекращаются, они только замедляются. Это объясняется тем, что при низких и отрицательных температурах вживой древесине должна сохраняться нормальная структура клетки. Проще говоря, живая клетка не должна замерзнуть, а для этого обменные процессы в древесине не должны прекращаться ни на минуту. Кроме того, в состоянии зимнего покоя в дереве постоянно происходит кутикулярная транспи-_ рация влаги с поверхности сбросивших листву веток и с поверхности ствола. И хотя интенсивность ее в 10—20 раз ниже листовой транспирации, все же объем испаренной влаги в состоянии зимнего покоя у деревьев лиственных пород может составить до 10—30 л в сутки. Увеличению зимней транспирации способствует низкая влажность морозной зимней атмосферы, а также иссушающее действие зимних ветров.
Помимо этих двух факторов, надо еще иметь в виду и то, что для сохранения своей жизнедеятельности живые клетки должны постоянно дышать. Под дыханием при этом подразумевается внутриклеточный процесс окисления органических веществ. Дыхание есть неотъемлемое свойство, которое присуще всем живым организмам, тканям и клеткам. Главным материалом для дыхания древесной клетки есть глюкоза. Во время дыхания в результате расщепления глюкозы, кроме углекислого газа и воды, выделяется 2867 кДж/моль (686 ккал/моль) энергии (Лесная энциклопедия, 1986). Эта энергия по большей части сохраняется в дереве в форме высокоэнергетических химических соединений, но часть освобождаемой энергии при этом выделяется в виде тепла. Таким образом, можно утверждать, что дерево в процессе своей жизнедеятельности как бы себя «подогревает». Зимой этот фактор благотворно влияет на процесс зимовки пчел в дупле.
Обобщая вышеизложенное, можно сказать, что в состоянии глубокого зимнего покоя в сбросивших листву деревьях обменные процессы в интересах обеспечения жизнедеятельности дерева хотя и замедляются, но полностью не прекращаются.
В соответствии с разработанной мною моделью функционирования живого дерева в зимнее время (см. приложение 1), проводящая система дерева продолжает выполнять свои функции осушения дупла с зимующим клубом пчел и в состоянии глубокого зимнего покоя при наступлении отрицательных температур. Однако в случае наступления продолжительного периода низких температур (ниже —15— 20 °С) дерево переходит в состояние вынужденного (экзогенного) покоя, при котором сокодвижение в стволе и ветвях прекращается. Чаще всего начало периода экзогенного покоя (в случае его наступления) приходится на декабрь— январь. Возобновление нормального сокодвижения в дереве в дальнейшем будет происходить в периоды продолжительных потеплений или же с началом весеннего сокодвижения в феврале-марте.
Таким образом, максимальная продолжительность периода вынужденного покоя в зависимости от географической широты может составлять в холодные зимы не более 1—3 месяцев. В южных широтах европейской части и в более северных широтах при мягкой зиме период экзогенного покоя может и вовсе не наступать. С точки зрения физиологической, для обеспечения нормальной жизнедеятельности дерева этот период (в отличие от периода органического покоя) не является обязательным. Просто под воздействием низких отрицательных температур дерево вынужденнопереходит в это состояние, хотя прекрасно может обходиться без периода экзогенного покоя в своем жизненном цикле.
А коль скоро этот период наступил, функционирование проводящей системы дерева в это время будет носить ограниченный характер и отвод влаги из дупла проводящей системой существенно уменьшится, а в отдельные периоды и вовсе прекратится. В основном в этот период влага из клуба при охлаждении отработанного воздуха будет оседать в нижнюю часть дупла, и накапливаться там, в т-рухлявой пористой ситовине. Поскольку плотность ситовины в 2—2,5 раза ниже, чем у живой древесины, а водопог-лощение и водопроницаемость выше (Уголев Б.Н., 1991), то в нижней незапрополисованной части дупла временно может аккумулироваться значительное количество влаги.
Если даже предположить, что в это время влага из дупла не будет удаляться другими путями, то все равно в нижней части дупла вся выдыхаемая пчелами влага (а это приблизительно по 1 кг влаги за каждый зимний месяц) сможет полностью разместиться в ситовине в виде сконденсированной воды, инея или льда. При этом содержащаяся в нижней холодной части дупла влага находится за пределами теплой обитаемой части и по этой причине не оказывает никаких вредных воздействий на зимний клуб. Однако при этом еще какая-то часть влаги будет удаляться из дупла вместе с отработанным воздухом при вентилировании пчелами гнездовой полости в активной фазе воздухообмена. Об этом более подробно поговорим чуть ниже.
Следует заметить, что в естественных- условиях законы живой природы в это тяжелое для пчел время все же «помогают», как говорится, чем могут. Известно, что, в соответствии с законами физики, при конденсации или замерзании влаги выделяется тепло. Так вот, при осуществлении этого процесса в нижней части дупла выделяемое «сухое» тепло за счет явления конвекции будет подниматься вверх к гнезду и подогревать клуб.
Следует обратить внимание и на такую особенность вла-гообменных процессов в дупле. Хорошо известно, что в теплые зимы пчелы, находящиеся в ульях, зимуют хуже, чем в холодные. Это связано прежде всего с наличием повышенной наружной и внутриульевой влажности. Что же касается дупла, то в теплые зимы в дереве период вынужденного покоя не наступает, и сокодвижение в нем будет осуществляться постоянно. Следовательно, на всем протяжении периода сложной зимовки в теплую зиму механизм поглощения влаги проводящей системой дерева будет постоянно отбирать из гнезда лишнюю влагу и максимально облегчать зимовку пчелам в дупле. В холодные зимы с началом создания в атмосфере условий для возобновления сокодвижения влага из ситовины будет удаляться проводящей системой дерева в первую очередь. Это объясняется тем, что, ввиду большой тепловой инерции почвенного слоя, корневое сокодвижение всегда будет возобновляться с задержкой по отношению к готовности дерева принимать влагу.
В летнее время, когда семья ведет активную жизнедеятельность, проблема удаления излишней влаги из дупла не является столь актуальной, как зимой. В этот период семья имеет возможность в полную силу использовать пчел-вентилировщиц для активной вентиляции гнезда. Помимо этого, как было сказано выше, удаление излишней влаги будет происходить также и за счет действия механизма удаления влаги проводящей системой живого дерева.
P.S. Когда я уже полностью закончил написание этого подраздела, мне удалось раздобыть подшивку журналов «Пчеловодство» за 1982 год, которую я раньше не читал, где в № 7 в статье «О вентиляции в ульях и зимовниках» Таранов Г.Ф. писал: «В дупле... пчелы тщательно замазывают прополисом потолок, а в летке оставляют небольшое отверстие. В таких условиях не может проходить вентиляция, достаточная для удаления водяных паров. В процесс удаления излишка воды включается само дерево. В дупле живого дерева в течение всей зимы жидкости протекают по сосудам снизу вверх. Даже во время морозов идет ток жидкости, хотя и замедленный. Пчелы пропитывают прополисом, непроницаемым для водяных паров, только потолок своего естественного жилья, оставляя открытыми боковые стенки дупла. Водяные пары проникают через них в сосуды дерева и уносятся в крону. Происходит медленное удаление воды. К тому же влага удаляется с углекислым газом, который оседает вниз дупла».
Мне кажется, что это высказывание из прошлого века лишний раз подтвердило обоснованность версии влагооб-менных процессов, о которой мы говорили выше.
♦ Версия воздухообменных процессов в дупле с пчелиным гнездом
Создаваемые в дупле естественным образом оптимальные условия для жизни пчел предполагают, в частности, наличие там оптимального воздухообмена. Это означает, что пчелы в дупле имеют возможность беспрепятственно получать необходимое для обеспечения их жизнедеятельности количество свежего воздуха (кислорода, О2) и удалять из гнезда отработанный воздух: углекислый газ (СО2) и водяной пар (Н2О). Попробуем разобраться, как происходят эти процессы. Однако сначала проведем краткое описание модели зимнего клуба, а точнее — тех его характеристик, которые будут непосредственно влиять на воздухообмен. Следует сказать, что в настоящее время существует довольно устоявшееся представление о структуре зимнего клуба, хотя некоторые авторы отдельные детали этой модели и могут трактовать по-разному.
Итак, зимний клуб пчел состоит из плотной, слабо проницаемой для воздуха, внешней корки плотно сидящих пчел. Толщина этой корки, в зависимости от внешней температуры, может изменяться от 1—2 до 5—7 см. Соответствующим образом изменяется и воздухопроницаемость корки — чем она толще, тем меньше ее воздухопроницаемость и наоборот. В корке клуба пчелы сидят упорядоченно: все они, плотно сцепившись, располагаются на сотах вверх головкой. При этом головка пчел, находящихся внизу, прячется под брюшком сидящих вверху пчел. Крылья пчел, расположенных в корке, полураскрыты и имеют возможность двигаться. Часть пчел корки клуба влезают в пустые ячейки сотов и сидит там совершенно неподвижно. Внутри под коркой располагается так называемый активный центр (ядро клуба), где пчелы могут свободно перемещаться, а воздух — свободно циркулировать. Плотность населения пчел в ядре клуба ниже, чем в корке. В каждой улочке клуба в одном месте сплошная корка имеет меньшую плотность и здесь образуется своеобразное отверстие (продух), через которое происходит вентиляция и отток отработанного воздуха. Казалось бы, что этот продух должен находиться вверху улочки. Однако, как показывает натурный эксперимент, в каждой улочке продух находится внизу (По-боженский Н.К., 1991). Скорее всего, так оно и есть по нескольким причинам.
Во-первых, в естественных условиях в дупле клуб в большинстве случаев находится выше летка и своим объемом перекрывает пустоту дупла (см. законы Ф.А. Соколова). Учитывая, что верхняя часть дупла герметична, удалять отработанный и получать свежий воздух клуб может только снизу.
Во-вторых, при необходимости зимний клуб может «включать» и активную вентиляцию гнездового пространства. Но поскольку пчелы в клубе располагаются вверх головками, то вентиляционный поток воздуха в этом случае тоже будет направлен вниз.
В-третьих, как показал Н. Решетников (2000), объем выдыхаемого пчелами отработанного воздуха на 30% больше объема потребляемого свежего воздуха. Это объясняется тем, что в процессе переработки меда к объему газовых составляющих выдыхаемого воздуха добавляется метаболическая вода в виде дополнительного объема водяного пара, который вырабатывается естественным образом в пчелином организме в результате окисления глюкозы меда.
Это утверждение о различии объемов вдыхаемого и выдыхаемого пчелами воздуха является совершенно новым взглядом на процесс дыхания у пчел и требует дальнейшего уточнения. Однако можно уже сейчас сказать, что косвенным подтверждением высказанному взгляду является тот факт, что дыхание у пчел несимметрично по времени — выдох обычно длится дольше, чем вдох (Таранов Г.Ф., 1968). Но, с другой стороны, А.Д. Комиссар (1994) утверждает, что в настоящее время пути и механизмы удаления метаболической воды из организма пчел в зимнем клубе практически не исследованы, и в свою очередь высказывает предположение, что одним из возможных путей удаления воды из организма пчел является выдыхание более теплого воздуха, содержащего большее количество паров воды.
Учитывая все эти аргументы, будем принимать утверждение Н. Решетникова в качестве рабочей гипотезы, которая требует дальнейшего подтверждения. Если же мы согласимся с этим утверждением, то зимний клуб надо будет рассматривать как своеобразный источник (генератор) небольшого избыточного давления. При этом избыточное давление будет постоянно «выталкивать» из клуба отработанный воздух вниз в сторону летка, ибо другого направления для его выхода в дупле просто нет.
Теперь возвратимся к непосредственному рассмотрению вопросов воздухообмена.
Известно, чтопри потреблении 1 кг меда пчелам для его усвоения необходимо получить 800 г кислорода, а выделяют они при этом 680 г водяного пара, 1000 г углекислого газа и около 12,57 МДж (3000 ккал) тепловой энергии.
Во время зимовки одна пчела съедает за сутки 2,5— 3,0 мг, в среднем — 2,75 мг меда. Тогда семья из 20 тыс. пчел за сутки потребляет 55 г меда и 44 г кислорода, а выделяет 38 г водяного пара и 55 г углекислого газа. Основные выделения пчелы делают с дыханием. Для того чтобы удалить из своего организма указанное количество Н2О и СО2, пчелы выдыхают воздух, имеющий 4% углекислого газа (нормальная атмосфера содержит 0,03% СО2) и почти 100%-ную влажность (Комиссар А.Д., 1994). Выходящий из клуба отработанный воздух подогрет и имеет температуру не ниже температуры корки клуба (8-10 °С), а содержание кислорода в нем может опускаться до 3—4% (в нормальной атмосфере содержится 21% О2).
Определив характеристики отработанного воздуха, рассмотрим сам механизм воздухообменных процессов в дуплес пчелиным клубом. Для начала будем считать, что объем вдыхаемого и выдыхаемого клубом воздуха одинаков.
До наступления холодной погоды, пока пчелы еще летают, воздухообмен в гнездовой полости осуществляется в основном за счет активной вентиляции через леток пчела-ми-вентилировщицами. После образования зимнего клуба воздухообмен дупла будет осуществляться еще и за счет других факторов, о которых и пойдет речь ниже.
Поскольку дупло представляет собой замкнутую полость с открытым отверстием (летком), то в принятых нами условиях холодный воздух, как более плотный и тяжелый, будет поступать («вливаться»)
Дата добавления: 2016-03-30; просмотров: 1981;