Электрическая активность путей и центров слухового анализатора
Для волокон слухового нерва даже в тишине характерна сравнительно высокая частота спонтанных разрядов (фоновой импульсации) —до 100 имп/с. При звуковом раздражении частота импульсации в волокнах нарастает в течение всего времени, пока действует звук. Степень учащения разрядов различна у разных волокон и связана с интенсивностью воздействия. Для каждого волокна слухового нерва может быть найдена так называемая оптимальная частота звука, дающая наибольшую частоту разрядов и наиболее низкий порог реакции. Эта оптимальная частота определяется местом на основной мембране, где расположены рецепторы, связанные с данным волокном. Таким образом, частотная избирательность волокон слухового нерва отражает пространственное кодирование информации в улитке, определяемое ее конструкцией.
Рисунок возбуждения нейронов центральных отделов слухового анализатора весьма разнообразен. Наиболее часто здесь встречаются клетки, возбуждение которых длится в течение всего времени действия стимулов, несколько снижаясь по частоте. Реже, чем в зрительном анализаторе, на низких уровнях в слуховом анализаторе встречаются нейроны, отвечающие лишь на включение и выключение звука или резкие перепады его интенсивности (оп-, оЦ-, оп—о/7-типа). На более высоких уровнях слухового анализатора процент таких нейронов возрастает. В слуховой зоне коры имеется много нейронов, вызванные разряды которых длятся десятки секунд после прекращения звука.
На каждом из уровней слухового анализатора с помощью макроэлектродов могут быть зарегистрированы характерные по форме вызванные потенциалы, отражающие синхронизированные реакции больших групп нейронов и волокон (рис. 224).
| он |

Рис.223. Импульсация волокон слухового нерва.
Схематическая иллюстрация принципа залпов (по Шварцкопфу). а—звуковой сигнал; б—и—реакции отдельных волокон; к—суммарная реакция этих волокон, повторяющая частоту звука.
Рис.224. Вызванные потенциалы разных уровней слуховой системы.
Суммарные ответы на звуковой щелчок в слуховом нерве (СН), кохлеарных ядрах (КЯ), верхней оливе (ВО), латеральной петле (ЛП), задних холмах четверохолмия (ЗХ), внутреннем коленчатом теле (ВКТ) и слуховой коре (СК). Отметка времени — 5 мс слева и 20 мс справа.
Анализ частоты звуков (высоты тонов)
Звуковые колебания вовлекают в колебательный процесс перилимфу верхнего и нижнего каналов улитки на всем их протяжении неодинаково. Колебательная реакция основной мембраны, передаваемая на эндолимфу, имеет характер бегущей волны, а локализация амплитудного максимума этой волны на мембране зависит от частоты звука. Таким образом, в процесс возбуждения вовлекаются разные клетки спирального (кортиева) органа, примыкающие к основной мембране.
Существенное значение для восприятия звуков разной частоты локализации рецеп-торных клеток спирального органа, вовлекаемых в возбуждение, доказывается, в частности, и поведенческими экспериментами. Если повредить у собаки завиток улитки у ее основания, то исчезают ранее выработанные условные рефлексы, на высокие тона;
если же повреждение нанести в области вершины улитки, исчезают условные рефлексы на низкие тона; разрушение только среднего завитка улитки вызывает выпадение тонов средней частоты диапазона. Раздельное пространственное представительство зон, возбуждаемых при действии звуков разной частоты, подтверждается электрофизиологическими методами (вызванные потенциалы, нейронная активность) на всех уровнях слухового анализатора:
Для сравнительно низких частот возможен анализ высоты тона за счет реакции группы волокон слухового нерва, повторяющей частоту действия раздражителя. Это означает, что существуют два сочетающихся механизма различения высоты тонов. При
действии высоких тонов происходит лишь пространственное кодирование, основанное на неодинаковом расположении возбужденных рецепторных клеток на основной мембране. При низких и средних тонах осуществляется и временное кодирование, когда информация передается по определенным группам волокон слухового нерва в виде импульсов, частота которых соответствует частоте воспринимаемых улиткой звуковых колебаний.
Основным отражением частотной настройки отдельных нейронов на всех уровнях слуховой системы является наличие у них так называемых частотно-пороговых показателей. Эти показатели отражают зависимость пороговой интенсивности звука, необходимой для возбуждения клетки, от его частоты. В обе стороны по диапазону частот от оптимальной, или характеристической, частоты порог реакции нейрона резко возрастает. Таким образом, элемент оказывается «настроенным» на выделение из всей совокупности звуков лишь определенного, достаточно узкого участка частотного-диапазона. Частотно-пороговые кривые разных клеток перекрывают весь частотный диапазон слышимых звуков. Форма этих кривых у многих нейронов высших слуховых центров значительно усложняется.
Анализ силы звуков (интенсивности звучания)
Сила звука кодируется числом возбужденных нейронов и частотой их импульсации. Увеличение числа возбужденных нейронов при действии все более громких звуков связано с тем, что нейроны слухового анализатора резко отличаются друг от друга по уровню возбудимости (порогам реакций). При слабых раздражителях в реакцию вовлекается лишь небольшое число наиболее чувствительных нейронов, а при усилении стимулов возбуждается все большее количество дополнительных нейронов. Кроме того, сами пороги возбуждения внутренних и наружных рецепторных клеток спирального органа неодинаковы. Возбуждение внутренних волосковых клеток возникает при большей силе звукового раздражения. Поэтому в зависимости от интенсивности звукового раздражения изменяется соотношение числа возбужденных внутренних и наружных волосковых клеток.
Звуковые ощущения
Тональность (частота) звука. Человек может воспринимать звуки с частотой колебания от 16 до 20 000 Гц. Этот диапазон соответствует 10—11 октавам. Верхняя граница воспринимаемых звуков зависит от возраста: чем человек старше, тем она ниже; старики часто не слышат высоких тонов (например, звука, издаваемого сверчком). У многих животных верхняя граница слуха лежит значительно выше: у собаки, например, удается получить условные рефлексы на очень высокие, неслышимые человеком звуки. Различение частоты звука характеризуется тем минимальным различием по частоте двух звуков, которое еще улавливается человеком. При низких и средних частотах человек способен заметить различия в 1—2 Гц. Встречаются люди с абсолютным слухом: они способны точно узнавать и обозначать любой звук даже при отсутствии звука сравнения.
Слуховая чувствительность. Минимальную силу звука, слышимого человеком в половине случаев его предъявления, называют абсолютной слуховой чувствительностью. Установлено, что пороги слышимости сильно изменяются в зависимости от частоты звука.
В области частот от 1000 до 4000 Гц слух человека обладает максимальной чувствительностью. В этих пределах слышен звук, имеющий ничтожную энергию порядка 1 • Ю"12 Вт/м2 (1 • Ю-9 эрг/с-см2). При звуках ниже 1000 и выше 4000 Гц чувствительность резко уменьшается: например, при 20 и при 20 000 Гц пороговая энергия звука должна быть около 1-10~3 Вт/м2 (1 эрг/с-см2) (нижняя кривая АЕГОО на рис. 225).
При увеличении силы звука неизменной частоты можно дойти догакой силы, когда звук вызывает неприятное ощущение давления и даже боли в ухе. Звуки такой силы
| Рис.225. Область звукового восприятия человека. Зависимость пороговой интенсивности звука (ось ординат — звуковое давление в дин/см2) от частоты тональных звуков (ось абсцисс в Гц). Линия АЕРОО абсолютные пороги, АВСО — пороги болевого ощущения от громких звуков. |

дадут, очевидно, верхний предел слышимости (кривая АВСО на рис. 225) и ограничат область слухового восприятия. Внутри этой области лежат и так называемые речевые поля, в пределах которых по частоте и интенсивности распределяются звуки речи.
Громкость звука. Кажущуюся громкость звука следует отличать от его физической силы. Ощущение громкости не идет строго параллельно нарастанию интенсивности звучания. Единицей громкости звука является бел. Эта единица представляет собой десятичный логарифм отношения действующей интенсивности звука / к пороговой его интенсивности /о. В практике обычно используется в качестве единицы громкости децибел (дБ), т. е. 0,1 бела, иначе говоря, 10 1§ю1/1о.
Нарастание ощущения громкости при усилении звука различно в зависимости от его частоты. В среднем диапазоне слышимых частот (1000 Гц) человек замечает изменение интенсивности всего на 0,59 дБ (5,9•10-2 Б), а на краях шкалы дифференциальный порог по громкости доходит до 3 дБ (0,3 Б). Максимальный уровень громкости, когда звук вызывает болевое ощущение, равняется 130—140 дБ над порогом слышимости человека. Таким образом, громкость звучания определяется сложным взаимодействием таких его показателей, как интенсивность (сила) и высота тона (частота).'
Адаптация. Если на ухо долго действует тот или иной звук, то чувствительность слуха падает. Степень этого снижения чувствительности (адаптация) зависит от длительности, силы звука и его частоты.
Механизмы адаптации в слуховом анализаторе изучены далеко не полностью. Известно, что сокращения т. 1епзог 1утрат и т. 51ареош5 могут изменять интенсивность звуковой энергии, передающейся на улитку. Кроме того, раздражение определенных точек ретикулярной формации среднего мозга приводит к угнетению вызванной звуком электрической активности кохлеарного ядра и слуховой зоны коры. Анатомическим образованием, через которое может опосредоваться это влияние на рецепторы, являются волокна, направляющиеся от ретикулярной формации к улитке и слуховым передаточным нейронам.
Бинауральный слух. Человек и животные обладают пространственным слухом, т. е. способностью определять положение источника звука в пространстве. Это свойство основано на наличии двух симметричных половин слухового анализатора (бинаураль-ный слух).
Острота бинаурального слуха у человека очень высока: он способен определять расположение источника звука с точностью порядка 1 углового градуса. Физиологической основой этого служит способность нейронных структур слухового анализатора оценивать интерауральные (межушные) различия звуковых стимулов по времени их прихода на каждое ухо и по их интенсивности. Если источник звука находится в стороне от средней линии головы, звуковая волна приходит на одно ухо несколько раньше и большей силы, чем на другое. Оценка удаленности звука от организма связана с ослаблением звука и изменением его тембра.
|
Бинауральный эффект часто изучают путем раздельной стимуляции правого и левого уха через наушники. При этом задержка между звуками уже в П-Ю^с (11 мкс) или различие в интенсивности двух звуков на 1 дБ, приводит к кажущемуся сдвигу локализации источника звука от средней линии в сторону более раннего или более сильного звука. На уровне задних холмов четверохолмия и в вышерасположенных отделах слухового анализатора найдены нейроны с острой специфической настройкой на определенный ограниченный диапазон интерауральных различий по времени и интенсивности. Найдены также клетки, реагирующие лишь на определенное направление движения источника звука в пространстве.
ВЕСТИБУЛЯРНЫЙ АНАЛИЗАТОР
Вестибулярная сенсорная система играет наряду со зрительным и кинестетическим анализаторами ведущую роль в пространственной ориентировке человека. Она передает и анализирует информацию об ускорениях или замедлениях, возникающих в процессе прямолинейного или вращательного движения, а также при изменении положения головы в пространстве. При равномерном движении или в условиях покоя рецепторы вестибулярного анализатора не возбуждаются. Импульсы от вестибулорецепторов вызывают перераспределение тонуса скелетной мускулатуры, что обеспечивает сохранение равновесия тела. Эти влияния осуществляются рефлекторным путем через ряд отделов ЦИС.
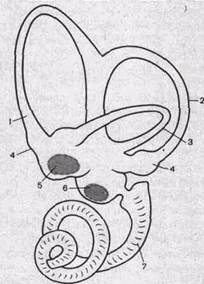
Периферическим отделом вестибулярного анализатора является вестибулярный аппарат, находящийся в лабиринте пирамиды височной кости. Он состоит из преддверия (уез1|Ьи1ит) и трех полукружных каналов (сапа1ез 5ет!с1гси1аг1з). Кроме вестибулярного аппарата в лабиринт входит улитка, в которой располагаются слуховые рецепторы. Полукружные каналы (рис. 226) располагаются в трех взаимно перпендикулярных плоскостях: верхний — во фронтальной, задний — в сагиттальной и наружный — в горизонтальной. Один из концов каждого канала расширен (ампула). Вестибулярный аппарат включает в себя также два мешочка: засси-1из и и<:пси1ив. Первый из них лежит ближе к улитке, а второй — к полукружным каналам. В мешочках преддверия находится отолитовый аппарат: скопления рецепторных клеток (вторичночувствующие ме-ханорецепторы) на возвышениях или пятнах (таси1а зассиП; таси1а иШсиН). Выступающая в полость мешочка часть рецепторной клетки оканчивается одним более длинным подвижным волоском и 60—80 склеенными неподвижными волосками. Эти волоски пронизывают желеобразную мембрану, содержащую кристаллики карбоната кальция — отолиты. Возбуждение волосковых клеток преддверия происходит вследствие скольжения отолитовой мембраны по волоскам, т. е. их сгибания (а не растяжения или сжатия, как считали раньше) (рис. 227).
| Рис.226. Строение лабиринта височной кости (схема). 1, 2. 3 — полукружные канальцы; 4 — ампулы каналов;5,6 — преддверие, которое разделяется на два мешочка; 7 — улитка. |
В перепончатых полукружных каналах, повторяющих форму костных каналов, заполненных, как и весь лабиринт, плотной эндолимфой (ее вязкость в 2—3 раза больше, чем у воды), рецепторные волос-ковые клетки сконцентрированы только в ампулах в виде крист (спз<;ае атри1апз). Они также снабжены волосками. При движении эндолимфы (во время угловых ускорений), когда волоски сгибаются в одну сторону, волосковые клетки возбуждаются, а при противоположно направленном движении — тормозятся. В волосковых клетках и преддверия, и ампулы при их сгибании генерируется рецепторный потен-
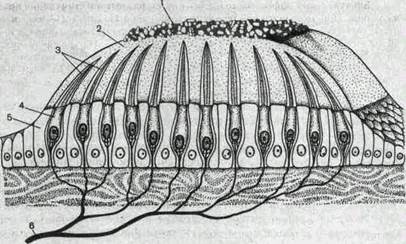
Рис.227. Строение отолитового аппарата.
1 — отолиты; 2 — отолитовая мембрана; 3 — волоски рецепторных клеток; 4 — рецепторные клетки; 5 — опорные клетки; (1— нервные во.шкна.
циал, который через синапсы (посредством выделения ацспихо.тна) передает сигналы о раздражении волосковых клеток окончаниям волокон вестибулярного нерва.
Волокна вестибулярного нерва (отростки биполярных нейронов) направляются в продолговатый мозг. Импульсы, приходящие по этим волокнам, поступают на нейроны бульбарного вестибулярного комплекса (ядра: преддверное верхнее Бехтерева, пред-дверное латеральное Дейтерса, Швальбе и др.). Отсюда сигналы направляются во многие отделы ЦНС: спинной мозг, мозжечок, глазодвигательные ядра, кору большого мозга, ретикулярную формацию и вегетативные ганглии.
Даже в полном покое в вестибулярном нерве регистрируется спонтанная импуль-сация. Частота разрядов в нерве повышается при поворотах головы в одну сторону и тормозится при поворотах в другую сторону (детекция направления движения). Реже частота разрядов повышается или, наоборот, тормозится при любом движении. Две трети волокон обнаруживают эффект адаптации (уменьшение частоты разрядов) во время действия углового ускорения.
Нейроны вестибулярных ядер обладают способностью реагировать и на изменение положения конечностей, повороты тела, сигналы от внутренних органов, т. е. осуществлять синтез информации, поступающей из разных источников. При этом они обеспечивают контроль и управление различными двигательными реакциями.
Важнейшие из этих реакций вестибулоспинальные, вестибуловегетативные и вести-булоглазодвигательные. Вестибулоспинальные влияния через вестибуло-, ретикуло-и руброспинальные тракты обеспечивают изменения импульсации нейронов сегментар-ных уровней спинного мозга. Таким образом осуществляются динамическое перераспределение тонуса скелетной мускулатуры и рефлекторные реакции, необходимые для сохранения равновесия. Мозжечок при этом ответствен за фазический характер этих реакций: после его удаления вестибулоспинальные влияния становятся по преимуществу тоническими. Во время произвольных движений вестибулярные влияния на спинной мозг ослабляются.
В вестибуловегетативные реакции вовлекаются сердечно-сосудистая система, желудочно-кишечный тракт и другие органы. При сильных и длительных нагрузках на вестибулярный аппарат возникает патологический симптомокомплекс, названный
болезнью движения (примером которой может служить морская болезнь). Последняя проявляется изменением сердечного ритма (учащение, а затем замедление), сужением, а затем расширением сосудов, усилением движения желудка, головокружением, тошнотой и рвотой. Повышенная склонность к болезни движения может быть уменьшена специальной тренировкой (вращение, качели) и применением ряда лекарственных средств.
Вестибулоглазрдвигательные рефлексы (глазной нистагм) состоят в ритмическом медленном движении глаз в противоположную вращению сторону, сменяющемся скачком глаз обратно. Само возникновение и характеристика вращательного глазного нистагма — важные показатели состояния вестибулярной системы и широко используются в авиационной, морской и космической медицине, а также в эксперименте и клинике.
Интересно, что при переходе от земных условий, когда отолитовый аппарат постоянно возбуждается ускорением силы тяжести, к условиям орбитальных полетов, характеризующихся невесомостью, деятельность отолитовых рецепторов изменяется: вначале (до 70 ч) их активность резко снижается, а затем увеличивается и лишь к 5 суткам возвращается к норме (адаптация). Это было выявлено путем регистрации импульсов в отдельных веточках вестибулярного нерва лягушек, запущенных в космос на специальном биоспутнике.
В коре больших полушарий мозга обезьян основные афферентные проекции вестибулярного аппарата локализованы в задней части постцентральной извилины. В моторной коре кпереди от нижней части центральной борозды обнаружена вторая вестибулярная зона. Исследование этих проекций осуществляется экспериментально путем электрического раздражения вестибулярного нерва и регистрации вызванных потенциалов в разных отделах мозга. Выяснено, что есть два основных пути поступления вестибулярных сигналов в кору обезьян: прямой — через дорсо-медиальную часть вентрального постлатерального ядра (УРЬ) и непрямой вестибуло-церебело-таламический путь через медиальную часть вентролатерального ядра (VI-).
Локализация вестибулярной зоны коры человека окончательно не выяснена. Вестибулярный анализатор помогает организму ориентироваться в пространстве при активном движении животного и при пассивном переносе с места на место с завязанными глазами. При этом лабиринтный аппарат с помощью корковых отделов системы анализирует и запоминает направление движения, повороты и пройденное расстояние. Следует подчеркнуть, что в нормальных условиях пространственная ориентировка обеспечивается совместной деятельностью зрительной и вестибулярной сенсорных систем.
Чувствительность вестибулярного анализатора здорового человека весьма высока: отолитовый аппарат позволяет воспринять ускорение прямолинейного движения, равное всего 2 см/с2. Порог различения наклона головы в сторону — всего около 1°, а вперед и назад — 1,5—2°. Рецеп-торная система полукружных каналов позволяет человеку замечать ускорения вращения в 2— 3° в 1 с2
Дата добавления: 2016-03-27; просмотров: 2051;