ОСОБЕННОСТИ КОДИРОВАНИЯ В АНАЛИЗАТОРАХ
Уже на уровне рецепторов осуществляется первичное кодирование стимулов: переход их из присущей им формы физической или химической энергии в форму нервных импульсов. Преобразованные таким образом сообщения поступают на следующий уровень анализаторной системы, где подвергаются дальнейшим преобразованиям, приводящим к изменению кода. Однако ни на одном из уровней' анализатора не происходит восстановление стимула в его первоначальной форме, т. е. декодирование. Этим физиологическое кодирование отличается от большинства технических систем связи, где сообщение, как правило, восстанавливается в первоначальном, декодированном виде.
Вторая принципиальная особенность нервного кодирования — множественность и перекрытие кодов. Это означает, что для одного и того же признака сигнала (например, его интенсивности) в анализаторе одновременно используется несколько различных вариантов нервных кодов: частота импульсации в отдельных нейронных каналах, число возбужденных элементов и их локализация («адрес») в нервном слое. Удельный вес каждого из этих кодов может изменяться на разных уровнях анализатора, но их параллельность сохраняется.
Еще одна особенность кодирования — это «зашумленность» большинства сенсорных кодов, т. е. добавление к импульсам, несущим информацию, фоновой импульсации. Это затрудняет анализ и восприятие информации.
Следует отметить еще две особенности нервного кодирования. Во-первых, скорости кодирования и перекодирования должны быть достаточно большими, чтобы избежать задержки реакций организма на принимаемый сигнал. Во-вторых, исследуемые нейро-физиологами нервные коды не наверняка являются кодами, используемыми мозгом. Методов, которые позволили бы точно установить, каким образом и какие именно нервные клетки получают и перекодируют то или иное сообщение, пока не существует. Сейчас трудно доказать, что именно тот принцип кодирования, который можно исследовать на данном уровне анализатора, используется нейронами следующего уровня. Поэтому можно говорить лишь о гипотетических нервных кодах.
Примеры отдельных видов кодирования. Код «начало — конец стимула» осуществляется в одиночном чувствительном элементе в основном на подкорковых уровнях анализатора и относится к временному кодированию. Его особенность состоит в том, что импульсация нейрона возникает только в момент включения, выключения или резкого изменения интенсивности раздражителя. Так, например, на всех уровнях зрительного анализатора описаны нейроны, отвечающие на включение света («оп-нейроны»), на его выключение («оН-нейроны»), либо же на включение и на выключение (оп-оП-нейроны). Такие реакции выделяют наиболее информативные временные контуры событий. Меньшему числу периферических чувствительных клеток свойственно кодирование параметров раздражителя с помощью частоты импульсации.
Для мозга очень важен код, характеризующийся изменением распределения импульсов внутри пачки или их последовательности. Импульсы могут быть по-разному сгруппированы в начале, середине или конце одной пачки импульсов, что придает ей разный временной рисунок. Для передачи сведений о сигнале имеют значение и изменения длительности пачки импульсов, число импульсов в пачке, степень колебаний частоты разрядов в их длительной последовательности. Важным кодовым признаком является последовательность включения отдельных нейронных каналов, по которым поступает информация о том, что раздражитель передвигается по рецепторной поверхности.
К кодам высокого уровня, функционирующим у высших позвоночных животных, как правило, в корковом отделе анализатора, относится кодирование посредством последовательного включения параллельно работающих каналов, изменением их числа, а также так называемое позиционное преобразование. Последний вид кодирования наиболее важен и заключается в том, что определенный признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в строго определенном месте того или иного нервного слоя. Например, возбуждение на корковом уровне зрительной системы определенной небольшой группы нейронов означает, что в определенной части поля зрения появилась световая полоска.определенного размера и ориентации.
На высших уровнях анализаторов происходит переход от преимущественно временного кодирования признаков раздражителя (свойственного периферическим отделам) к преимущественно пространственному (в основном позиционному) коду.
V. Детектирование сигналов — специальный вид избирательного анализа отдельных признаков раздражителя и их конкретного биологического значения. Осуществляют такой анализ специализированные нейроны-детекторы, которые благодаря свойствам своих связей способны реагировать лишь на строго определенные параметры стимула.
Корковые зрительные детекторы реагируют лишь на одно из множества положений или наклонов светлой или темной полоски, расположенной в определенной части поля зрения. При другом положении той же полоски ответят другие нейроны. Совокупность нейронов, оценивающих разные стороны одного и того же признака (например, все возможные ориентации изображений), составляет систему детекции этого признака.
Общим в распределении детекторов является иерархический принцип, согласно которому на более низких уровнях локализуются детекторы более простых признаков, обеспечивающие простой анализ. В высших отделах анализатора, как правило, сконцентрированы детекторы более сложных признаков.
VI. Опознание образов -- конечная и наиболее сложная операция анализатора. Она заключается в классификации образа, отнесении его к тому или иному классу объектов, с которыми ранее встречался организм. Это происходит на основе всей предыдущей обработки афферентного сигнала, после расщепления его нейронами-детекторами на отдельные признаки и их раздельного параллельного анализа. Задача операции опознания может быть сведена к построению мозгом «модели раздражителя» и ее выделению из множества других подобных моделей. Опознание завершается принятием решения о том, с каким объектом или ситуацией встретился организм. Полагают, что для этого существуют специфические пространственно связанные наборы нейронов (нейронные ансамбли—высшие детекторы), возбуждение которых означает для мозга появление того или иного образа. Именно в результате такого опознания мы осознаем, какого человека видим перед собой, чей голос слышим, какой запах чувствуем и т. д.
Опознание происходит независимо от изменчивости сигнала. Мы надежно опознаем, например, предметы при различной их освещенности, окраске, размере, ракурсе, ориентации и положении в поле зрения. Точно так же знакомый голос опознается при разной его громкости, наличии звукового фона, а смысл речи — и при значительных изменениях ее тембра и темпа. Отсюда следует, что на каких-то высших уровнях анализатора организуется независимое от этих изменений признаков отражение сигнала —сенсорныи обр.чз. Это совокупность сигналов, отображаемых в сходном пространственно-временном распределении процессов возбуждения и торможения на высшем уровне анализатора.
Взаимодействие нейронов анализаторов осуществляется с помощью двух основных механизмов — возбудительного и тормозного. Возбудительное взаимодействие представлено главным образом между элементами последовательных нервных слоев, в то время как тормозное — в основном между нейронами одного и того же слоя.
Возбудительное взаимодействие организуется следующими способами. Во-первых, аксон каждого нейрона, приходя в вышележащий слой, делится на большее или меньшее число веточек, вступающих в синаптическне контакты не с одним, а с несколькими нейронами.
Во-вторых, «дендритное дерево», т. е. входы нейрона, имеют, как правило, синапти-ческие контакты с аксонами не одной, а нескольких клеток предыдущего слоя. Из-за этого практически все нейроны анализатора имеют проекционные поля, т. е. совокупность нейронов на следующем и более высоких уровнях анализатора, с которыми они взаимодействуют.
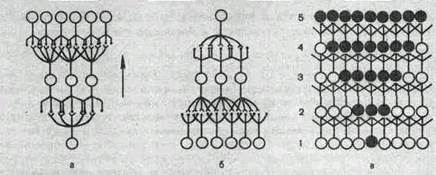
Рис.209. Схема проекционного (а) и рецептивного (б) полей нейрона. Нервная сеть (в). Направление потока импульсов показано стрелкой. Возбужденные нейроны в последовательных слоях (1—5) выделены.
Совокупность рецепторов, импульсы от которых поступают на данный нейрон, называют его рецептивным полем.
Следует иметь в виду, что рецептивные и проекционные поля существуют у всех нейронов системы одновременно. Более того, они частично перекрываются. Такое «сдвинутое боковое перекрытие» связей заключается в том, что часть рецепторов, входящих в рецептивное поле данного нейрона, входит и в рецептивное поле соседней с ним клетки, а часть нейронов, входящих в проекционное поле какого-либо рецептора, может входить и в проекционное поле соседнего рецептора (рис. 209).
Такое сложное взаимодействие клеток приводит к образованию в анализаторе так называемой нервной сети. Благодаря ей происходит повышение чувствительности анализаторов к слабым сигналам. Кроме того, избыточность связей в сетевых структурах обеспечивает анализаторам высокую приспособляемость к меняющимся условиям среды.
Тормозное взаимодействие в анализаторах осуществляется с помощью тормозных вставочных нейронов (интернейронов). Чаще всего торможение заключается в том, что каждый возбужденный нейрон активирует тормозной интернейрон, который в свою очередь подавляет, затормаживает импульсацию как самого возбудившего его элемента, так и его соседей по слою. Сила этого торможения тем больше, чем сильнее возбужден элемент и чем ближе к нему соседняя клетка. За счет такого универсального механизма торможения в анализаторных системах осуществляется значительная часть операций по снижению избыточности информации, поступающей из рецепторов, а также по выделению сведений о пространственных и временных признаках раздражителей.
Торможение играет ведущую роль и во временной обработке сигналов, так как включается, как правило, с некоторой задержкой после возбуждения и подавляет преимущественно поздние по отношению к началу раздражения ответы нейронов. В результате этого они хорошо отвечают только на изменения раздражителя во времени, в частности на его включение и выключение.
Дата добавления: 2016-03-27; просмотров: 2915;