Функции фибробластов
1. Продукция всех компонентов межклеточного вещества (волокон и основного аморфного вещества). Фибробласты синтезируют коллаген, эластин, фибронектин, гликозаминогликаны и др.
2. Поддержание структурной организации и химического гомеостаза межклеточного вещества (за счет сбалансированных процессов его выработки и разрушения).
3. Регуляция деятельности других клеток соединительных тканей и влияние на другие ткани. Продукция цитокинов (колониестимулирующих факторов гранулоцитов и макрофагов).
4.Заживление ран. При воспалении и заживлении ран фибробласты активируются макрофагами.
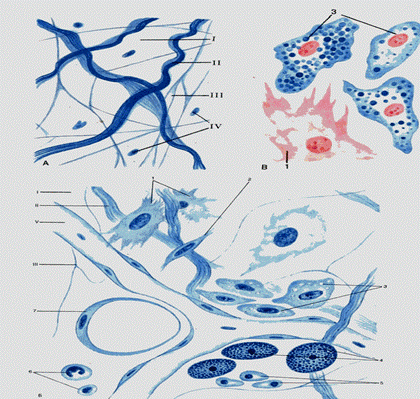 Рис. 3.2. Рыхлая и волокнистая соединительные ткани – пленочный препарат I – основное вещество; II – коллагеновые волокна; III – эластические волокна; IV – клетки; V – кровеносный сосуд. 1 – фибробласты, 2 – фиброцит, 3 – макрофаги, 4 – тучные клетки, 5 – плазмоциты, 6 – лейкоциты, 7 – жировая клетка.
Рис. 3.2. Рыхлая и волокнистая соединительные ткани – пленочный препарат I – основное вещество; II – коллагеновые волокна; III – эластические волокна; IV – клетки; V – кровеносный сосуд. 1 – фибробласты, 2 – фиброцит, 3 – макрофаги, 4 – тучные клетки, 5 – плазмоциты, 6 – лейкоциты, 7 – жировая клетка.
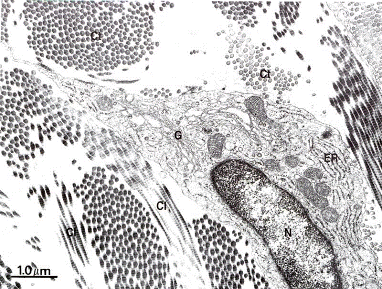
Рис.3.3. Электронограмма фибробласта среди коллагеновых волокон
(х 18.500).
Ct- поперечные,
Сl – продольные срезы коллагеновых волокон;
N – ядро клетки смещено на периферию;
ER – эндоплазматический ретикулум;
G – комплекс Гольджи.
|
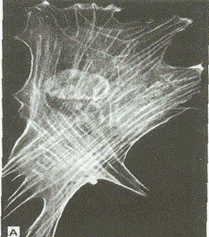
Рис. 3.4. Актиновые микрофиламенты в цитоплазме миофибробласта (иммунофлюоресцентный метод).
Макрофаги. На втором месте в количественном отношении среди клеток рыхлой соединительной ткани стоят макрофаги Макрофаги образуются путем дифференцировки и размножения, вышедших в ткань из крови моноцитов. Различают свободные и фиксированные макрофаги.По сравнению с фибробластами они меньших размеров 10-15 мкм. Имеют различную форму - округлую, вытянутую или неправильную. В базофильной цитоплазме макрофагов содержится много лизосом, фагосом, пиноцитозных пузырьков. Умеренное развитие имеют митохондрии, ЭПС, комплекс Гольджи. Макрофаги – активно фагоцитирующие клетки, богатые органеллами для внутриклеточного переваривания поглощенного материала (лизосомы) и синтеза антибактериальных и других биологически активных веществ (пироген, антиферон, лизоцим, ЭПС). Ядра содержат больше хроматина и окрашиваются более интенсивно, чем ядра фибробластов. Цитоплазма макрофагов образует глубокие складки и длинные микроворсинки, которые обеспечивают захват инородных частиц. Поверхность макрофага имеет рецепторы чувствительные к эритроцитам, T и B-лимфоцитам, антигенам и иммуноглобулинам. Последние обеспечивают возможность их участия в иммунных реакциях организма.
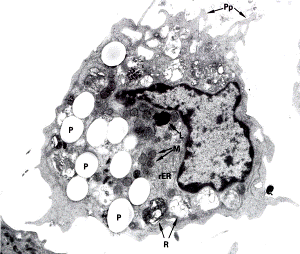 А А
|  Б Б
|
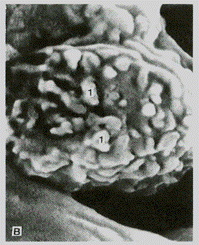
Рис. 3.5. Ультраструктура макрофага. А – активная форма, Б –поверхность макрофага (х11.600). Сканирующая электронная микроскопия. 1– отростки клетки. Pp, 1 –псевдоподии; Р –фагоцитированные частицы; М – митохондрии; L – лизосомы. Ядро неправильной формы.
Макрофаги наряду со способностью к фагоцитозу синтезируют целый ряд веществ, обеспечивающих врожденный иммунитет (лизоцим, интерферон, пироген и др.). Макрофаги секретируют медиаторы - монокины, способствующие специфической реакции на антигены и цитолитические факторы, которые избирательно разрушают опухолевые клетки.
Функции макрофагов:
1. фагоцитоз: распознавание, поглощение и переваривание поврежденных, зараженных, опухолевых и погибших клеток, компонентов межклеточного вещества, а также экзогенных материалов и микроорганизмов.
2. участие в индукции иммунных реакций, т.к. (играют роль антиген-представляющих клеток).
3. регуляция деятельности клеток, других типов (фибробластов, лимфоцитов, тучных клеток, эндотелиоцитов и др.).
Макрофаги развиваются из моноцитов. Совокупность клеток, имеющих одно ядро, называется монокулиарной фагоцитарной системой, и мононукледов, обладающих способностью к фагоцитозу: захватывать из тканевой жидкости организма инородные частицы, погибающие клетки, неклеточные структуры, бактерии и др. Фагоцитированный материал подвергается внутри клетки ферментативному расщеплению (“завершенный фагоцитоз”), благодаря чему ликвидируются вредные для организма агенты, возникающие местно или проникающие извне. Макрофаги (гистиоциты) рыхлой волокнистой соединительной ткани, звездчатые клетки синусоидных сосудов печени, свободные и фиксированные макрофаги кроветворных органов (костного мозга, селезенки, лимфатических узлов), макрофаги легкого, воспалительных экссудатов (перитонеальные макрофаги), остеокласты, гигантские клетки инородных тел и глиальные макрофаги нервной ткани (микроглия). Все они способны к активному фагоцитозу, имеют на своей поверхности рецепторы к иммуноглобулинам и происходят из промоноцитов костного мозга и моноцитов крови. В отличие от таких “профессиональных” фагоцитов способность к факультативному поглощению может быть выражена независимо от указанных циторецепторов у других клеток (фибробласты, ретикулярные клетки, эндотелиоциты, нейтрофильные лейкоциты). Но эти клетки не входят в состав макрофагической системы.
И.И. Мечников (1845-1916) первым пришел к мысли о том, что фагоцитоз, возникающий в эволюции как форма внутриклеточного пищеварения и закрепившийся за многими клетками, одновременно является важным защитным механизмом. Он обосновал целесообразность объединения их в одну систему и предложил назвать ее макрофагической. Макрофагическая система представляет собой мощный защитный аппарат, принимающий участие, как в общих, так и в местных защитных реакциях организма. В целостном организме макрофагическая система регулируется как местными механизмами, так нервной и эндокринной системами. В 30-40-х годах эту защитную систему называли ретикулоэндотелиальной. В последнее время ее называют системой мононуклеарных фагоцитов, что, однако, неточно характеризует ее в связи с тем, что среди клеток, входящих в эту систему, есть и многоядерные (остеокласты).
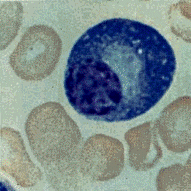
Плазматические клетки – плазмоциты имеют округлую форму. Величина плазматических клеток от 7 до 10мкм. Ядро округлой или овальной формы лежит, как правило, эксцентрично. Глыбки хроматина в нем расположены по радиусам. Они напоминают пирамиды, основание которых лежит на ядерной оболочке. Создается впечатление, что хроматин расположен в виде спиц в колесе. Данное обстоятельство служит одним из диагностических признаков при определении плазмоцитов.
 А А
|  Б Б
|  В В
|
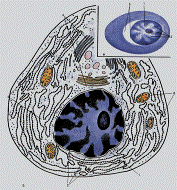
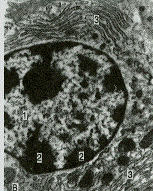
Рис. 3.6. Плазматическая клетка. А – в мазке крови. Б – схема. В – электронограмма.
Цитоплазма клеток резко базофильна, особенно по периферии. В центре перед ядром имеется небольшое просветление - "дворик". Он содержит сетчатый аппарат, центриоли, митохондрии. Цитохимически в плазматических клетках обнаруживается громадное количество рибонуклиопротеидов, обусловливающих базофилию цитоплазмы. Среди белков обнаруживается много – γ-глобулина. С ним связывается основная функция клеток - участие в защитных реакциях организма.
Зрелые плазматические клетки характеризуются высокой базофилией и эксцентрично расположенным ядром. Под электронным микроскопом определяются параллельные мембраны. Наличие параллельных мембран в цитоплазматической сети характерно для клеток, синтезирующих белок на “экспорт”. Вырабатываемый плазматической клеткой белок может иметь различный состав и определяется качеством белка раздражителя или антигена. Поэтому мы говорим, что синтез белка в плазматических клетках - частное выражение способности этих клеток принимать участие в белковом обмене. Наряду с этим цитоплазма клетки выделяет небольшое количество гликозаминогликанов, поступающих в межклеточное вещество.
Сравнение концентрации глобулина показало, что в зрелых клетках его меньше, чем в незрелых. В последнее время считают, что зрелая клетка - это плазматическая клетка в состоянии покоя. При встрече с антигеном, раздражителем она также может интенсивно образовывать глобулин и по своим морфологическим признакам приближаться к той клетке, которую называют "незрелой". Плазматические клетки называют иммунокомпетентными, т. к. они сохраняют "память" об антигенных раздражителях и при повторной встрече с ним блокируют антиген специфическим антителом.
Одно из проявлений иммунной реакции у позвоночных животных при попадании в организм чужеродного агента - выделение плазматическими клетками антител.
В цитоплазме плазматических клеток могут появляться кристаллические включения, воспринимающие кислые красители, так называемые тельца Русселя. Считают, что они являются конгломератами глобулинов, синтезированных ранее этой клеткой.
Плазматические клетки обеспечивают гуморальный иммунитет путем выработки антител. За 1 секунду каждый плазмоцит синтезирует до нескольких тысяч молекул иммуноглобулииов (более 10 млн. молекул в час).
Тканевые базофилы (лаброциты, тучные клетки). Тучные клетки – постоянный клеточный компонент рыхлой волокнистой соединительной ткани, осуществляющий важные регуляторные функции. Эти клетки имеют в цитоплазме зернистость, напоминающую гранулы базофильных лейкоцитов. Они являются регуляторами местного гомеостаза соединительной ткани.
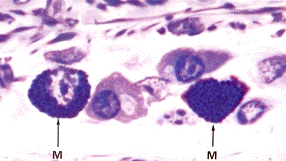 А А
|  Б Б
|
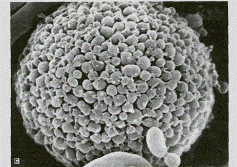
Рис. 3.7. Структура тучной клетки А – Тучные клетки (М) в составе соединительной ткани (х1200); Б – рельеф клеточной поверхности.
Развитие тучных клеток осуществляется в тканях из предшественника, который имеет, как предполагают, костномозговое происхождение. На их дифференцировку и рост влияют факторы клеточного микроокружения (фибробласты, эпителиальные клетки и их продукты). В отличие от базофилов, которые после миграции в ткани живут недолго (от нескольких часов до нескольких суток), тучные клетки обладают сравнительно большой продолжительностью жизни (от нескольких недель до нескольких месяцев). В течение этого периода под действием соответствующих стимулов тучные клетки, очевидно, способны делиться.
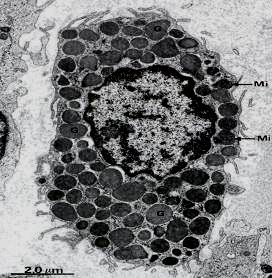
Рис. 3.8. Электронограмма тучной клетки (х12.000). G – крупные гранулы заполняют всю цитоплазму; Мi – митрхондрии расположенные между ними, в центре расположено ядро.
Тканевые базофилы имеют разнообразную форму. У человека и млекопитающих чаще их форма овальная. Размеры 3,5х14 мкм. Ядро небольшое, богатое хроматином. Встречаются двуядерные клетки.
Гранулы тучных клеток содержат разнообразные биологически активные вещества. Субмикроскопически они представляют плотные тельца неправильной формы диаметром 0,3-1,4 мкм, окрашиваются метахроматично. Клетки содержат митохондрии, внутриклеточный сетчатый аппарат. Компоненты тучных клеток у различных животных и в различных участках соединительной ткани различные. У кроликов и морских свинок тучных клеток мало, у белых мышей очень много. У человека и животных тучные клетки обнаружены во всех местах, где имеются прослойки рыхлой соединительной ткани. Они расположены группами по ходу кровеносных и лимфатических сосудов. Количество тучных клеток изменяется при различных состояниях организма - при беременности увеличивается количество тучных клеток в матке и молочных железах, в желудке и кишечнике в разгар пищеварения. Тучные клетки содержат разнообразные медиаторы и ферменты.
Структурно-функциональные различия тучных клеток. Популяция тучных клеток образована элементами, которые обладают неодинаковыми морфофункциональными свойствами и могут качественно и количественно различаться даже в пределах одного органа. Высказывают предположение о том, что отдельные субпопопуляции тучных клеток выполняют в организме неодинаковые функции.
Функции тучных клеток:
1. Гомеостатическая, которая осуществляется в физиологических условиях путем медленного выделения небольших количеств биологически активных веществ, способных влиять на различные тканевые функции – в первую очередь, на проницаемость и тонус сосудов, поддержание баланса жидкостей в тканях.
2. Защитная и регуляторная, которая обеспечивается путем локального выделения медиаторов воспаления и хемотаксических факторов, обеспечивающих (а) мобилизацию эозинофилов и различных эффекторных клеток, участвующих в так называемых реакциях поздней фазы; (б) воздействие на рост и созревание соединительной ткани в зоне воспаления.
3. Участие в развитии аллергических реакций вследствие наличия высокоаффинных рецепторов к иммуноглобулинам класса Е (IgE) на их плазмолемме и функциональной связи этих рецепторов с секреторным механизмом. Участие тучных клеток в развитии аллергических реакций, как и базофильных гранулоцитов включает:
Ø связывание IgE с высокоаффинными рецепторами на их плазмолемме;
Ø взаимодействие мембранного IgE с аллергеном;
Ø активацию и дегрануляцию тучных клеток с выделением содержащихсяв их гранулах веществ и продукцией ряда новых.
Ø предполагается, что тучные клетки выполняют магниторецепторную функцию.
Дегрануляция может опосредоваться также рецепторами комплемента или вызываться белками нейтрофилов, протеиназами, нейропептидами (вещество Р, соматостатин), лимфокинами.
По подсчетам Уокера полная смена тучных клеток рыхлой соединительной ткани может произойти за 16 – 18 месяцев. По данным Н.Г.Хрущева за 9 дней.
Таблица 3.2.
Медиаторы и ферменты, содержащиеся в тучных клеток
| Медиатор | Функция | |
| Гистамин | Н1, Н2 – рецептор опосредованное действие на гладкомышечные клетки (ГМК), эндотелий, нервные волокна. Вазодилатация, повышение проницаемости капилляров, отек, хемокинез, бронхоспазм, стимуляция афферентных нервов | |
| Химаза | Расщепление коллагена IV типа, глюкагона, нейротензина, фибронектина | |
| Триптаза | Конверсия С3 в С3а, расщепление фибриногена, фибронектина, активация коллагеназы | |
| Карбоксипептидаза В | Разборка внеклеточного матрикса | |
| Дипептидаза | Конверсия LTD4 в LTE4. Разрушение внеклеточного матрикса | |
| Кининогеназа | Конверсия кининогена в брадикинин | |
| Инактиватор фактора Хагемана | Инактивация фактора Хагемана | |
| Гексозаминидаза, глюкуронидаза, галактозидаза | Разрушение внеклеточного матрикса (гликопротеинов, протеогликанов) | |
| β-Гликозаминидаза | Расщепление гликозаминов | |
| Пероксидаза | Конверсия Н2О2 в Н2О, инактивация лейкотриенов, образование липидных пероксидов | |
| Фактор хемотаксиса эозинофилов (ECF) | Хемотаксис эозинофилов | |
| Фактор хемотаксиса нейтрофилов (NCF) | Хемотаксис нейтрофилов | |
| Гепарин | Антикоагулянт, избирательно связывает антитромбин III. Ингибитор альтернативного пути активации комплемента. Модифицирует активность других ранее синтезированных медиаторов. | |
| Простагландин PGD2, тромбоксан TXA2 | Сокращение ГМК бронхов, вазодилатация, увеличение сосудистой проницаемости, агрегация тромбоцитов | |
| Лейкотриены LTC4, LTD4, LTE4, медленно реагирующий фактор анафилаксии SRS-A | Вазо- и бронхоконстрикция, увеличение сосудистой проницаемости, отеки. Хемотаксис и /или хемокинез |
Жировые клетки, липоциты. Различают две разновидности жировых клеток: клетки белого и бурого жира. Клетки белого жира моновакуалярные, имеют одну жировую вакуоль. Они располагаются в рыхлой соединительной ткани главным образом по ходу сосудов, а в некоторых участках организма (под кожей, между лопатками, в сальнике и других местах) образуя значительные скопления. Это позволяет выделить специальную жировую ткань, построенную почти исключительно из жировых клеток. Жировые клетки имеют шарообразную форму. Они больше по размеру других клеток соединительной ткани. Их диаметр 30-50 мкм. Непосредственными предшественниками жировых клеток являются малодифференцированные соединительнотканные клетки, расположенные главным образом около капилляров (перикапиллярные или адвентициальные клетки). Возможно образование липоцитов из гистиоцитов, фагоцитирующих жировые капли. В процессе дифференцировки в жировой клетке накапливаются мелкие капли нейтрального жира, которые путем слияния образуют более крупные. Основная функция липоцитов - запас жира как макроэргического соединения. При распаде его высвобождается большое количество энергии, используемой организмом как источник тепла, а также для фосфорилирования АДФ с образованием АТФ. Жир служит источником образования воды, выполняет защитную и опорную функцию. Жировые клетки синтезируют биологически активные вещества – лептин, регулирующий чувство насыщения, эстрогены и т.п.
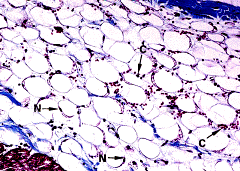 А А
|  Б Б
|
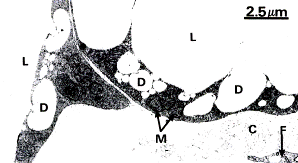
Рис.3.9. Клетки белого жира (апудоциты, моновакуолярные клетки) А- совокупность жировых клеток образует жировую дольку, снабженную большим количеством кровеносных сосудов (С) х480); Б – электронная микрофотография периферии 2-х апудоцитов, L – жировая вакуоль; D – мелкие капельки жира; М- митохондрии; С-коллагеновые волокна в межклеточном пространстве. (х6.000).
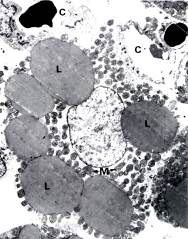
Рис. 3.10. Электронная микрофотография клетки бурого жира: Ядро расположено в центре,
L – жировые вакуоли,
М- митохондрии,
С – капилляры.
Жировые клетки кроме роли энергетического депо выполняют функции эндокринной железы, гормоны которой регулируют объем и массу тела. Этим гормоном является лептин.
Белая жировая ткань составляет 15-20 % массы тела взрослых самцов и на 5 % больше у самок. В некотором смысле о ней можно говорить как о крупном метаболически активном органе, поскольку она участвует главным образом в поглощении из крови, синтезе, хранении и мобилизации нейтральных липидов (жиров). (Мобилизовать жир – значит сделать его подвижным, с тем чтобы использовать как горючее” в других частях тела.) В жировой клетке при температуре тела жир находится в состоянии жидкого масла. Он состоит из триглицеридов содержащих три молекулы жирной кислоты, образующие эфир с глицерином. Триглицериды – наиболее калорийный вид питательных веществ, поэтому жир в жировых клетках представляет собой хранилище „высококалорийного” горючего, притом относительно легкого. Кроме того, у обитателей холодных стан жир участвует в регуляции температуры лежащих под ним органов. И, наконец, жир служит отличным заполнителем различных „щелей” в организме и образует „подушки”, на которых могут лежать те или иные внутренние органы.
Бурые жировые клетки обнаружены у новорожденных детей и у некоторых животных на шее, около лопаток, за грудиной, вдоль позвоночника, под кожей между мышцами. Она состоит из жировых клеток, густо оплетенных гемокапиллярами. Клетки бурого жира -поливакуолярные. Диаметр клеток бурого жира почти в 10 раз меньше, чем диаметр клеток белого жира. Эти клетки принимают участие в процессах теплопродукции. Адипоциты бурой жировой ткани имеют множество мелких жировых включений в цитоплазме. По сравнению с клетками белой жировой ткани здесь обнаруживается множество митохондрий. Бурый цвет жировым клеткам придают железосодержащие пигменты – цитохромы митохондрий. Окислительная способность бурых жировых клеток примерно в 20 раз выше белых и почти в 2 раза превышает окислительную способность мышцы сердца. При понижении температуры окружающей среды активность окислительных процессов в бурой жировой ткани повышается. При этом выделяется тепловая энергия, обогревающая кровь в кровеносных капиллярах. В регуляции теплообмена определенную роль играет симпатическая нервная система и гормоны мозгового вещества надпочечников – адреналин и норадреналин, который через циклический аденозинмонофосфат стимулирует активность тканевой липазы, расщепляющей триглицериды на глицерин и жирные кислоты. Последние, накапливаясь в клетке, разобщают процессы окислительного фосфорилирования, что приводит к высвобождению тепловой энергии, обогревающей кровь, протекающую в многочисленных капиллярах между липоцитами. При голодании бурая жировая ткань изменяется меньше, чем белая.
Пигментоциты (пигментные клетки).содержат в своей цитоплазме пигмент меланин. Они имеют отростчатую форму и подразделяются на два вида - меланоциты, которые вырабатывают пигмент, и – меланофоры, способные лишь накапливать его в цитоплазме. У людей черной и желтой рас пигментные клетки более распространены, чем определяется неизменяемый в зависимости от времени года цвет кожи. Пигментоциты имеют короткие непостоянной формы отростки. Эти клетки лишь формально относятся к соединительной ткани, так как располагаются в ней. В настоящее время имеются веские доказательства того, что эти клетки образуются из нервных гребней, а не из мезенхимы.
Таблица 3.3. Различия между белой и бурой жировыми клетками
| Белая жировая клетка | Бурая жировая клетка |
| Широко распространена у человека: в т.ч. находится - в подкожной жировой клетчатке, - в сальнике, - в жировых отложениях вокруг внутренних органов, - в диафизах трубчатых костей (жёлтый костный мозг) и т.д. | а) Встречается у новорождённых детей - в области лопаток, - за грудиной и в некоторых других местах. б) У взрослого человека находится в воротах почек и в корнях лёгких. У животных, впадающих в спячку |
| В клетках ядра оттеснены к периферии. | Ядра расположены в центре клеток. |
| В клетках - одна большая жировая капля. | В клетках - много мелких жировых капель. |
| Количество митохондрий невелико. | В цитоплазме - много митохондрий (откуда - бурый цвет ткани). |
| Функции клетки: депонирование жира, ограничение теплопотерь, механическая защита. | Функция - обеспечение теплопродукции. |
| жир из белой жировой клетки расходуется, главным образом, не в ней самой, а в иных органах и тканях, | а жир бурой жировой клетки расщепляется для обеспечения теплопродукции непосредственно в ней самой. |
Адвентициальные клетки. Это малоспециализированные клетки, сопровождающие кровеносные сосуды. Они имеют уплощенную или веретенообразную форму со слабобазофильной цитоплазмой, овальным ядром и слаборазвитыми органеллами. В процессе дифференцировки эти клетки, по-видимому, могут превращаться в фибробласты, миофибробласты, адипоциты. Многие авторы отрицают существование адвентициальных клеток как самостоятельного клеточного типа, считая их клетками фибробластического ряда.
Эндотелиальные клетки – выстилают сосуды, поэтому их совокупность называется сосудистым эндотелием. Строение сосудистого эндотелия сходно со строением эпителиальной ткани. Эндотелию присущи следующие общие признаки.
1. Пограничное положение покровного эпителия и эндотелия.
2. Непрерывность эндотелиальной выстилки внутри всех кровеносных и лимфоносных сосудов у позвоночных.
3. Отсутствие основного промежуточного вещества по всей окружности клеток эндотелия и эпителия.
4. Наличие базальной мембраны, выполняющей функцию опоры и фиксации эндотелиальных клеток. Её основу, как и основу базальных мембран эпителия, составляет коллаген IV типа.
5. Гетерополярность в строении клеток. У эндотелиоцитов это проявляется в образовании микроворсинок на люминальной поверхности клеток (при относительной гладкости базальной), в неравнозначности элементов цитоскелета и концентрации микропиноцитозных везикул в цитоплазме противостоящих поверхностей клеток.
6. Специализированные контакты между эндотелиальными клетками по типу замыкающих, фибриллярные полоски которых располагаются ближе к люминальной поверхности клеток, чем подчеркивается её полярность.
7. Барьерная, секреторная, транспортная функции в их идеальном сочетании.
8. Рост эндотелия в тканевых культурах в виде монослоя клеток полигональной формы, обладающих выраженным контактным торможением.
В силу этого сходства многие исследователи относят эндотелий к эпителиальной ткани. Однако эндотелий происходит из мезенхимы, на основании чего его относят к соединительной ткани.
Эндотелиальные клетки играют важную роль в процессах транскапиллярного обмена, принимают участие в образовании тканевых мукополисахаридов, гистамина, фибринолитических факторов.
Функции эндотелия:
1. Транспортная – через него осуществляется избирательный двусторонний транспорт веществ между кровью и другими тканями. Механизмы: диффузия, везикулярный транспорт (с возможным метаболическим превращением транспортируемых молекул).
2. Гемостатическая – играет ключевую роль в свертывании крови. В норме образует атромбогенную поверхность; вырабатывает прокоагулянты (тканевый фактор, ингибитор плазминогена) и антикоагулянты (активатор плазминогена, простациклин).
3. Вазомоторная – участвует в регуляции сосудистого тонуса: выделяет сосудосуживающие (эндотелин) и сосудорасширяющее (простациклин, эндотелиальный релаксирующий фактор – окись азота) вещества; участвует в обмене вазоактивных веществ – ангаотензина, норадреналина, брадикинина.
4. Рецепторная – экспрессирует на плазмолемме ряд соединений, обеспечивающих адгезию и, и последующую трансэндотелиальную миграцию лимфоцитов, моноцитов и гранулоцитов.
5. Секреторная – вырабатывает митогены, ингибиторы и факторы роста, цитокины, регулирующие кроветворение, пролиферацию и дифференцировку Т- и В-лимфоцитов, привлекающие лейкоциты в очаг воспаления.
6. Сосудообразовательная – обеспечивает новообразование капилляров (ангиогенез) – как в эмбриональном развитии, так и при регенерации.
Перициты– клетки звездчатой формы, примыкающие снаружи к артериолам, венулам и капиллярам. Наиболее многочисленны в посткапиллярных венулах. Имеют собственную базальную мембрану, сливающуюся с базальной мембраной эндотелия, так что создается впечатление, что перицит заключен в расслоившуюся базальную мембрану эндотелия. Перицит охватывает стенку сосуда, что позволяет предположить их участие в регуляции просвета сосудов.
Перициты имеют дисковидное ядро с небольшими углублениями, содержат обычный набор органелл, мультивезикулярные тельца, микротрубочки и гликоген. В области, обращенной к стенке сосуда, содержат пузырьки. Около ядра и в отростках присутствуют сократительные белки, в т.ч. актин и миозин. Перициты покрыты базальной мембраной, но тесно связаны с эндотелиальной клеткой, т.к. базальная мембрана между ними может и отсутствовать. В этих местах выявлены щелевые и адгезионные контакты.
Функции перицитов четко не установлены. О конкретных функциях можно говорить с разной степенью вероятности.
1. Контрактильные свойства. Вероятно участие перицитов в регуляции просвета микрососуда.
2. Источник гладкомышечных клеток (ГМК). При заживлении ран и восстановлении сосудов перициты в течение 3-5 дней дифференцируются в ГМК.
3. 3.Влияние на эндотелиальные клетки. Перициты контролируют пролиферацию эндотелиальных клеток, как при нормальном росте сосудов, так и при их регенерации; модулируют функцию эндотелиальных клеток, регулируя транспорт макромолекул из капилляров в ткани.
4. Секреторная функция. Синтез компонентов базальной мембраны капилляра.
5. Участие в фагоцитозе.
Межклеточное вещество рыхлой волокнистой соединительной ткани состоит из волокон и основного аморфного вещества. Оно является продуктом деятельности клеток этой ткани, в первую очередь, фибробластов.
Функции межклеточного вещества рыхлой волокнистой соединительной ткани:
1.обеспечение архитектоники, физико-химических и механических свойств ткани;
2.участие в создании оптимального микроокружения для деятельности клеток;
3.объединение в единую систему всех клеток соединительной ткани и обеспечение передачи информации между ними;
4.воздействие на многочисленные функции различных клеток (пролиферацию, дифференцировку, подвижность, экспрессию рецепторов, синтетическую и секреторную активность, чувствительность к действию различных стимулирующих, ингибирующих и повреждающих факторов и т.п.). Этот эффект может осуществляться путем контактного воздействия компонентов межклеточного вещества на клетки, а также благодаря его способности накапливать и выделять факторы роста.
Коллагеновые волокна в составе разных видов соединительной ткани определяют их прочность. В рыхлой неоформленной волокнистой соединительной ткани они располагаются в различных направлениях в виде волнообразно изогнутых, спиралевидно скрученных, округлых или уплощенных в сечении тяжей толщиной 1-3 мкм и более. Длина их различна. Внутренняя структура коллагенового волокна определяется фибриллярным белком — коллагеном, который синтезируется на рибосомах гранулярной эндоплазматической сети фибробластов.
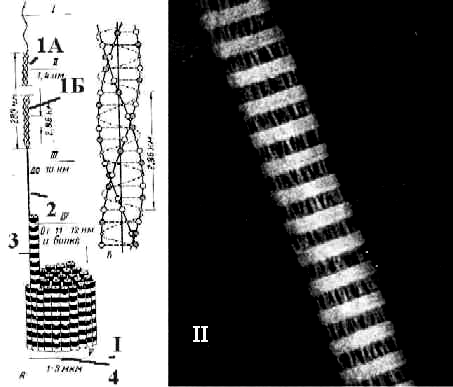
Рис. 3.11. I. Схема – уровни структурной организации коллагеновых волокон. II. Электронная микрофотография - коллагеновая фибрилла. Различают четыре уровня организации коллагеновых волокон: молекулы тропоколлагена (1), протофибриллы (2), фибриллы (3) и волокна (4).\
Коллагеновые волокна распространены не только в собственно соединительной ткани, но также в кости и хряще, где они соответственно называются оссеиновыми и хондриновыми. Эти волокна определяют прочность тканей на разрыв. В рыхлой неоформленной соединительной ткани они располагаются в различных направлениях в виде волнообразно изогнутых тяжей толщиной 1-3 мкм. Коллагеновые волокна состоят из пучков параллельно расположенных микрофибрилл толщиной в среднем 50-100 нм, связанных между собой гликозаминогликанами и протеогликанами. Их толщина зависит от числа фибрилл, которые имеют поперечную исчерченность (черные и светлые участки) с периодом повторяемости 64-70 нм. В пределах одного периода находятся вторичные полосы шириной 3-4 нм.
Коллагеновые структуры, входящие в состав соединительных тканей организма человека и животных, являются наиболее распространенными ее компонентами. Основным их компонентом является волокнистый белок - коллаген.
Коллаген - главный белок соединительной ткани, которая составляет свыше 50% веса организма человека и животных. Одновременно, по расчетам швейцарского ученого Ф. Верцара, на долю коллагена приходится около 30% общего количества белка в организме. Следовательно, коллаген в количественном отношении стоит среди белков на первом месте.
Расшифровка первичной структуры коллагена - важнейший этап развития этих знаний. Значение раскрытия структуры коллагена следует расценивать с учетом того большого интереса, который проявляют к коллагену в различных областях знаний. Он лежит в основе целых областей технологии. Все кожевенное производство - это по существу переработка коллагена. Денатурированный коллаген–желатин незаменимый компонент фото-киноматериалов. Из переработанного коллагена изготовляется множество материалов, применяющихся в ветеринарной и медицинской практике.
Экстрагированные из волокон молекулы коллагена имеют длину 200 нм и ширину 1,4нм. Они получили название тропоколлагена. Молекулы построены из трипластов - трех полипептидных цепочек, которые сливаются в единую спираль. Каждая цепочка содержит набор из трех аминокислот, закономерно повторяющихся на протяжении ее длины. Первая кислота в таком наборе может быть любой, вторая - пролин или лизин, третья – глицин.
Расположение аминокислот может варьировать, вследствие чего образуется четыре типа коллагена.
1 тип - в собственно соединительной ткани, кости, роговице глаза, склере, зубной связке и др.
2 тип - в гиалиновом и фиброзном хряще, стекловидном теле.
3 тип - в дерме кожи плода, кровеносных сосудах, в ретикулярных волокнах.
4 тип - в базальных мембранах, в капсуле хрусталика.
В 1973 году была расшифрована одна из полипептидных цепей коллагена, что представляется выдающимся событием. Коллаген значительно крупнее по молекулярному весу, чем другие изученные белки. Трудности работы по установлению структуры коллагена были обусловлены величиной молекулы и особой монотонностью его строения - частотой повторения аминокислотных остатков и их сочетаний, что сильно осложняло задачу исследований.
Молекулы коллагена имеют длину около 280 нм и ширину 1,4 нм. Они построены из триплетов — трех полипептидных цепочек, предшественника коллагена – проколлагена, свивающихся еще в клетке в единую спираль. Это первый, молекулярный, уровень организации коллагенового волокна. Проколлаген секретируется в межклеточное вещество.
Второй, надмолекулярный, уровень — внеклеточной организации коллагенового волокна — представляет агрегированные в длину и поперечно связанные с помощью водородных связей молекулы тропоколлагена, образующиеся путем отщепления концевых пептидов проколлагена. Сначала образуются протофибриллы, а 5-6 протофибрилл, скрепленных между собой боковыми связями, составляют микрофибриллы толщиной около 5 нм.
При участии гликозаминогликанов, также секретируемых фибробластами, формируется третий, фибриллярный и, уровень организации коллагенового волокна. Коллагеновые фибриллы представляют собой поперечно исчерченные структуры толщиной в среднем 20-100 нм. Период повторяемости темных и светлых участков 64-67 нм. Каждая молекула коллагена в параллельных рядах, как полагают, смещена относительно соседней цепи на четверть длины, что служит причиной чередования темных и светлых полос. В темных полосах под электронным микроскопом видны вторичные тонкие поперечные линии, обусловленные расположением полярных аминокислот в молекулах коллагена.
Четвертый, волоконный, уровень организации. Коллагеновое волокно, образующееся путем агрегации фибрилл, имеет толщину 1-10 мкм (в зависимости от топографии). В него входит различное количество фибрилл – от единичных до нескольких десятков. Волокна могут складываться в пучки толщиной до 150 мкм.
Коллагеновые волокна отличаются малой растяжимостью и большой прочностью на разрыв. В воде толщина сухожилия в результате набухания увеличивается на 50%, а в разбавленных кислотах и щелочах – в 10 раз, но при этом волокно укорачивается на 30%. Способность к набуханию больше выражена у молодых волокон. При термической обработке в воде коллагеновые волокна образуют клейкое вещество (греч. коllа — клей), что и дало название этим волокнам.
Ретикулярные (ретикулиновые, аргирофильные) волокна. Они встречаются в рыхлой и некоторых других видах соединительной ткани, в строме кроветворных органов, печени, внутренних оболочках сосудов. На препаратах импрегнированных серебром они располагаются в виде сети.
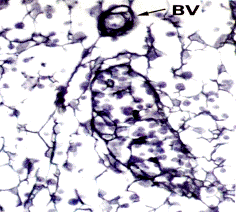
Рис. 3.12. Ретикулярные волокна в лимфатическом узле при импрегнации азотнокислым серебром. Волокна ветвятся, образуя тонкую сеть. ВV- кровеносный сосуд (х800).
Вопрос о природе ретикулярных волокон остается спорным. Большинство исследователей считают, что ретикулин - белок, составляющий основу этих волокон, представляет вещество близкое к коллагену, а импрегнационные и гистохимические отличия ретикулярных волокон от коллагеновых связаны со свойствами прошивающих волокна гликозаминогликанов. В отличие от коллагена и эластина ретикулин содержит больше серина, оксилизина и глютаминовой кислоты.
Эластические волокна. Эластические волокна придают ткани эластичность. Они менее прочны, чем коллагеновые на разрыв. В рыхлой соединительной ткани они образуют петлистую сеть, анастомозируя друг с другом. Толщина волокон от 0,2 до 1 мкм. В отличии от коллагеновых - они не имеют микроскопически видимых фибрилл и субмикроскопической поперечной исчерченности.
 А
А
|  Б
Б
|
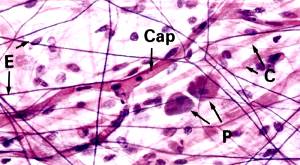
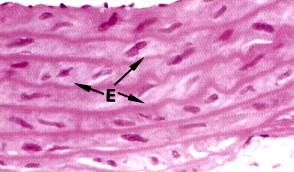
Рис. 3.13. А - Эластические волокна в соединительной ткани (х320). Б - эластические волокна в стенке крупной артерии (х400), Е – тонкие эластические волокна, Сар - разветвленный капилляр, Р –плазматические клетки, С- коллагеновые волокна.
Основой эластических волокон является глобулярный гликопротеин — эластин, синтезируемый фибробластами и гладкими мышечными клетками (первый, молекулярный, уровень организации). Для эластина характерны большое содержание пролина и глицина и наличие двух производных аминокислот – десмозина и изодесмозина, которые участвуют в стабилизации молекулярной структуры эластина и придании ему способности к растяжению, эластичности. Молекулы эластина, имеющие глобулы диаметром 2,8 им, вне клетки соединяются в цепочки — эластиновые протофибриллы толщиной 3-3,5 нм (второй, надмолекулярный, уровень организации). Эластиновые протофибриллы в сочетании с гликопротеином (фибриллином) образуют микрофибриллы толщиной 8-19 нм (третий, фибриллярный, уровень организации). Четвертый уровень организации – волоконный. Наиболее зрелые эластические волокна содержат около 90% аморфного компонента эластических белков (эластина) в центре, а по периферии – микрофибриллы. В эластических волокнах в отличие от коллагеновых нет структур с поперечной исчерченностью на их протяжении.
Дата добавления: 2016-02-16; просмотров: 6767;