Отношения организмов в биоценозах
Основу возникновения и существования биоценозов представляют отношения организмов, их связи, в которые они вступают друг с другом, населяя один и тот же биотоп. Эти связи определяют основные условия жизни видов в сообществе, возможности добывания пищи и завоевания нового пространства.
Классификации биоценотических отношений могут строиться с использованием разных принципов. Один из популярных подходов – оценка возможного результата контактов двух особей. Для каждой из них результат принимается как положительный, отрицательный или нейтральный. Сочетания результатов по 2 из 3-х возможных дают формальную схему из 6 вариантов, которая и положена в основу этой классификации.
7.3.1. Отношения хищник-жертва, паразит-хозяин
Отношения типа хищник-жертва, паразит-хозяин – это прямые пищевые связи, которые для одного из партнеров имеют отрицательные, а для другого – положительные последствия. По существу, к этому типу экологических взаимодействий можно отнести все варианты пищевых связей. Хотя пасущуюся на лугу корову и дятла, добывающего личинок из-под коры дерева, обычно не называют хищниками, тот тип взаимоотношений, в которые они вступают с организмами, служащими им пищей, имеет много общего с отношениями хищника и его жертв. Поэтому в западной литературе связь хищник– жертва понимается обычно в широком смысле, включая все формы добывания пищи. Они, в свою очередь, подразделяются на несколько категорий: 1) истинное хищничество, или хищничество в узком смысле слова; 2) паразитизм; 3) собирательство и 4) пастьба.
Хищниками обычно называют животных, питающихся другими животными, которых они ловят и умерщвляют. Для хищников характерно специальное охотничье поведение.
Добыча жертвы требует от них значительных затрат энергии на поиск, погоню, захват, преодоление сопротивления жертв.
Если размеры жертв намного меньше размеров питающихся ими животных, численность объектов питания высока и сами они легкодоступны – в этом случае деятельность плотоядного вида превращается в поиск и простой сбор добычи и называется собирательство.
Собирательство требует затрат энергии в основном на поиск, а не на захват пищи. Такое «собирание» характерно, например, для ряда насекомоядных птиц – куликов-зуйков, ржанок, зябликов, коньков и др. Однако между типичным хищничеством и типичным собирательством у плотоядных существует множество промежуточных способов добывания пищи. Например, ряд насекомоядных птиц характеризуется охотничьим поведением при поимке насекомых (стрижи, ласточки). Сорокопуты, мухоловки подстерегают и затем нагоняют жертву как типичные хищники. С другой стороны, способ питания плотоядных собирателей очень похож на собирание неподвижной пищи растительноядными животными, например семеноядными птицами или грызунами (горлица, сизый голубь, чечевица, лесная мышь, хомяки и др.), для которых также характерны специализированные поисковые формы поведения.
К собирательству можно отнести фильтрационное питание водных животных, седиментацию, или осаждение водной взвеси, сбор пищи илоедами или дождевыми червями. К нему же примыкает так называемое хищничество растений. У многих растений при недостатке азота в питании развиты способы улавливания и фиксации прилетающих к ним насекомых и переваривание белков их тел протеолитическими ферментами (пузырчатки, росянки, непентесы, венерина мухоловка и др.).
По способу овладения пищевыми объектами собирательство приближается к типичной пастьбе фитофагов. Специфика пастьбы заключается в поедании неподвижного корма, находящегося в относительном изобилии, на поиски которого не приходится тратить много усилий. С экологической точки зрения такой способ питания характерен как для стада копытных на лугу, так и для листогрызущих гусениц в кроне дерева или личинок божьих коровок в колониях тлей.
Таким образом, несмотря на экологическую специфичность разных способов питания, они связаны между собой всевозможными переходами, а иногда встречаются вместе у одного и того же вида. Такие же переходы можно встретить в природе между типичным хищничеством и паразитизмом.
Паразитизм такая форма связей между видами, при которой организм-потребитель использует живого хозяина не только как источник пищи, но и как место постоянного или временного обитания (рис. 85). По существу, типичный паразитический характер имеют связи насекомых-вредителей с растениями. Паразиты обычно намного мельче своего хозяина.

Рис. 85. Паразиты певчих птиц
Во взаимоотношениях хищник-жертва, паразит-хозяин наиболее ярко проявляется эволюционная и экологическая роль пищевых связей организмов. Хищничество, связанное с активным поиском и энергичными способами овладения сопротивляющейся и убегающей добычей, ведет к выработке разнообразных экологических адаптаций как у жертв, так и у их потребителей. При активном способе защиты от врагов естественный отбор способствует развитию у жертв органов чувств, быстроты реакции, скорости бега, инстинктов обманного поведения, что сопровождается совершенствованием нервной системы и ведет к прогрессивной эволюции группы.
При пассивном способе защиты развиваются покровительственная окраска, твердые панцири, шипы, иглы, инстинкты затаивания, использования недоступных хищникам убежищ и т. п. Некоторые из этих способов защиты характерны не только для малоподвижных или сидячих видов, но и для активно спасающихся от врагов животных.
Защитные адаптации у потенциальных жертв весьма разнообразны, иногда очень сложны и неожиданны. Например, каракатицы, спасаясь от преследующего хищника, опорожняют свой чернильный мешок. По гидродинамическим законам жидкость, выброшенная из мешка быстро плывущим животным, некоторое время не расплывается, приобретая форму обтекаемого тела, по размерам близкого к самой каракатице. Обманутый темным контуром, оказавшимся перед глазами, хищник «хватает» чернильную жидкость, наркотизирующее действие которой на время вообще лишает его возможности ориентироваться в окружающей среде. Своеобразен способ защиты у рыб-иглобрюхов. Их укороченное тело покрыто прилегающими шипами. Большой мешок, отходящий от желудка, позволяет этим рыбам в случае опасности раздуваться в шар, заглатывая воду; при этом их иглы расправляются и делают животное практически неуязвимым для хищника. Попытка крупной рыбы схватить иглобрюха может кончиться для нее смертью от застрявшего в глотке колючего шара.
В свою очередь, трудность обнаружения и поимки жертв способствует у хищников отбору на лучшее развитие органов чувств (зоркость, тонкий слух, чутье и т. д.), на более быструю реакцию на добычу, выносливость при преследовании и т. п. Таким образом, экологические связи хищников и жертв направляют ход эволюции сопряженных видов.
Хищники обладают обычно широким спектром питания. Добыча жертв требует много сил и энергии. Специализация ставила бы хищников в сильную зависимость от численности определенного вида жертв. Поэтому большинство видов, ведущих хищный образ жизни, способно переключаться с одной добычи на другую, особенно на ту, которая в данный период более доступна и многочисленна. Правда, у многих хищников есть предпочитаемые виды жертв, которых они добывают чаще, чем других. Эта избирательность может быть обусловлена разными причинами. Во-первых, хищник активно выбирает наиболее полноценную в кормовом отношении пищу. Например, нырковые утки и сиговые рыбы в северных водоемах выбирают среди водных беспозвоночных преимущественно личинок комаров-хирономид (мотыль), и их желудки иногда бывают наполнены мотылем, несмотря на наличие в водоеме и другого корма.
Характер пищи может быть обусловлен также пассивной избирательностью: хищник в первую очередь поедает такие корма, к добыче которых он наиболее приспособлен. Так, многие воробьиные питаются всеми насекомыми, обитающими открыто на поверхности почвы, на траве, листьях и т. п., но не поедают почвенных беспозвоночных, для добычи которых нужны специальные приспособления. Наконец, третьей причиной пищевой избирательности хищников может быть активное переключение на наиболее массовую добычу, появление которой стимулирует охотничье поведение. При высокой численности леммингов даже соколы-сапсаны, основной способ охоты которых – добывание птиц в воздухе, начинают охотиться на леммингов, схватывая их с земли. Способность переключения с одного вида добычи на другой – одно из необходимых экологических приспособлений в жизни хищников.
Паразитизм, в отличие от хищничества, характеризуется более узкой специализацией видов. Поскольку хозяин обеспечивает паразиту не только пищу, но и микроклимат, защиту и т. п., то чем лучше приспособленность паразита к особенностям организма хозяина, тем вероятнее его успех в размножении и оставлении потомства.
Тесная связь паразита с хозяином имеет следствием отбор двоякого рода. Среди паразитов получают преимущество те, которые способны более полно и длительно использовать хозяина, не приводя его к слишком ранней гибели и обеспечивая тем самым себе наилучшее существование. Иными словами, паразит изнуряет, но не губит хозяина. В свою очередь, отбор на сопротивляемость организма хозяина также приводит к тому, что вред от присутствия в нем паразита становится все менее и менее ощутимым. В ходе эволюции первоначально острые отношения хозяина и паразита могут перейти в нейтральные, а порой даже во взаимополезную постоянную связь двух видов. Примером такой смягченной историческим ходом развития системы паразит – хозяин могут служить трипаносомы в крови африканских антилоп, которые не приносят ощутимого вреда этим животным. В то же время у человека после передачи ему трипаносом переносчиком – мухой-цеце развивается смертельная «сонная болезнь». Показано, что умеренное объедание листвы насекомыми стимулирует ростовые процессы, в результате чего растения более полно развивают фотосинтетический аппарат, таким образом, некоторая степень «вреда» от насекомых-фитофагов даже полезна растительности.
Катастрофический вред от паразитов выражен преимущественно лишь в тех связях, которые еще не стабилизированы длительным ходом естественного отбора. Поэтому случайно завезенные вредители поражают сельскохозяйственные растения или животных часто намного сильнее, чем местные.
Среди многообразных форм паразитических отношений есть и такие, при которых гибель хозяина – обязательное следствие пребывания в нем паразита. Этот тип связей особенно распространен у насекомых, откладывающих свои яйца в яйца или личинки других. Такие насекомые получили название паразитоидов. Гибель хозяина обусловлена малым запасом в нем пищи, которой едва хватает на развитие одной или немногих личинок вида-потребителя.
Паразитизм, таким образом, связан всевозможными переходами с другими типами взаимоотношений.
Основная экологическая роль хищничества, паразитизма и других вариантов пищевых связей в сообществах заключается в том, что, последовательно питаясь друг другом, живые организмы создают условия для круговорота веществ, без которого невозможна жизнь. Вторая не менее важная роль этих отношений – взаимная регуляция численности видов.
7.3.2. Комменсализм
Комменсализм – это такая форма взаимоотношений между двумя видами, когда деятельность одного из них доставляет пищу или убежище другому (комменсалу). Иными словами, комменсализм – одностороннее использование одного вида другим без принесения ему вреда. Комменсализм, основанный на потреблении остатков пищи хозяев, называют еще нахлебничеством. Таковы, например, взаимоотношения львов и гиен, подбирающих остатки недоеденной львами добычи. Комменсалами крупных акул являются сопровождающие их рыбыприлипалы и т. д. Отношение нахлебничества устанавливается даже между насекомыми и некоторыми растениями. В жидкости кувшинов насекомоядных непентесов обитают личинки стрекоз, защищенные от переваривающего действия ферментов растения. Они питаются насекомыми, которые попадают в ловчие кувшины. Потребители экскрементов также комменсалы других видов.
Особенно развито использование убежищ либо в постройках, либо в телах других видов. Такой комменсализм называется квартирантством. Рыбки Fieraster прячутся в водяных легких голотурий, молодь других рыб – под зонтиками защищенных стрекательными нитями медуз. Комменсализмом является поселение растений-эпифитов на коре деревьев. В гнездах птиц, норах грызунов обитает огромное количество видов членистоногих, использующих микроклимат убежищ и находящих там пищу за счет разлагающихся органических остатков или других видов сожителей. Многие виды специализированы на таком образе жизни и вне нор не встречаются совсем. Постоянные норовые или гнездовые сожители получили название нидиколов.
Отношения типа комменсализма очень важны в природе, так как способствуют более тесному сожительству видов, более полному освоению среды и использованию пищевых ресурсов.
Нередко, однако, комменсализм переходит в другие типы отношений. Например, в гнездах муравьев среди большого числа их сожителей встречаются виды жуков-стафилинид из родов Lomechusa и Atemeles. Их яйца, личинки и куколки содержатся вместе с молодью муравьев, которые ухаживают за ними, облизывают и переносят в специальные камеры. Взрослых жуков муравьи также кормят. Однако жуки и их личинки поедают яйца и личинок хозяев, не встречая отпора с их стороны. На боках груди и первых сегментах брюшка у этих жуков имеются особые выросты – трихомы, у основания которых выделяются капельки секрета, чрезвычайно привлекающего муравьев. Секрет содержит эфиры, оказывающие на муравьев одурманивающее, наркотизирующее воздействие, подобное влиянию алкоголя. Муравьи постоянно облизывают ломехуз и атемелесов. В результате у них расстраиваются инстинкты, нарушается координация движений и появляются даже некоторые морфологические изменения. Рабочие муравьи в семьях, где много ломехуз, малоподвижные и вялые. Семьи становятся малочисленными и в результате погибают.
7.3.3. Мутуализм
В природе широко распространены взаимовыгодные отношения видов, называемые мутуализм. Мутуалистические связи могут возникать на основе предшествующего паразитизма или комменсализма. Степень развития взаимовыгодного сожительства может быть самой различной – от временных, необязательных контактов до такого состояния, когда присутствие партнера становится обязательным условием жизни каждого из них. Такие неразделимые полезные связи двух видов получили название симбиоза.
Классический пример симбиотических отношений – лишайники, представляющие тесное сожительство гриба и водоросли. В состав лишайников входят представители трех классов грибов – аскомицетов, базидиомицетов и фикомицетов. В свободном состоянии лишайниковые грибы, по-видимому, не встречаются в природе. Среди водорослей обнаружены представители 28 родов из отделов сине-зеленых, желто-зеленых, зеленых и бурых. Большинство их встречается и в свободноживущем состоянии. Симбиоз, по всей вероятности, возник из паразитизма гриба на водорослях. Гифы гриба, оплетая клетки и нити водорослей, образуют специальные всасывающие отростки – гаустории, проникающие внутрь протопласта через стенки клеток. Через них гриб получает вещества, ассимилированные водорослями. Воду и минеральные вещества водоросли получают из гиф гриба. Гриб постепенно убивает клетки водорослей, а затем использует их остатки, переходя к сапрофитному питанию. Однако степень паразитизма гриба умеренна. В лишайниках всегда лишь часть водорослевых клеток поражена грибными гифами, остальные продолжают успешно расти и развиваться. Замечено также, что гифы внедряются лишь в клетки, достигшие стадии зрелости. Эти особенности можно рассматривать как адаптацию для обеспечения размножения водорослевых компонентов лишайников. Всего в природе насчитывается более 20 000 видов этих симбиотических организмов, что говорит об успехе такого способа существования.
Типичный симбиоз представляют отношения термитов и их кишечных сожителей – жгутиковых отряда Hypermastigina. Эти простейшие вырабатывают фермент b-глюкозидазу, переводящий клетчатку в сахара. Термиты не имеют собственных кишечных ферментов для переваривания целлюлозы и без симбионтов погибают от голода. Появившиеся из яиц молодые термиты облизывают анальные отверстия взрослых, заражая себя жгутиконосцами. Жгутиковые находят в кишечниках термитов благоприятный микроклимат, защиту, пищу и условия для размножения. В свободноживущем состоянии они фактически не встречаются в природе.
Кишечные симбионты, участвующие в переработке грубых растительных кормов, обнаружены у многих животных: жвачных, грызунов, жуков-точильщиков, личинок майских жуков и др. Виды, питающиеся кровью высших животных (клещи, пиявки и др.), как правило, имеют симбионтов, помогающих переваривать ее.
У многоклеточных животных и растений симбиоз с микроорганизмами распространен очень широко. Известно сожительство многих видов деревьев с микоризными грибами, бобовых растений – с клубеньковыми бактериями Rhizobium, фиксирующими молекулярный азот воздуха. Симбионты-азотфиксаторы обнаружены на корнях около 200 видов других групп покрытосеменных и голосеменных растений. Симбиоз с микроорганизмами заходит иногда так далеко, что колонии симбиотических бактерий можно рассматривать как специализированные органы многоклеточных. Таковы, например, мицетомы каракатиц и некоторых кальмаров – мешки, наполненные светящимися бактериями и входящие в состав органов свечения – фотофоров.
Грань между симбиозом и иными типами отношений иногда весьма условна. Интересно использование своей кишечной микрофлоры зайцеобразными и некоторыми грызунами. У кроликов, зайцев, пищух обнаружено регулярное поедание собственных фекалий. Кролики производят два типа экскрементов: сухие и мягкие, покрытые слизистой оболочкой. Мягкие фекалии они слизывают прямо с ануса и проглатывают не разжевывая. Исследования показали, что такая копрофагия вполне естественна. Кролики, лишенные возможности потреблять мягкий кал, худеют или плохо прибавляют в массе и чаще подвержены различным заболеваниям. Мягкий кал кроликов – это почти неизмененное содержимое слепой кишки, обогащенное витаминами (преимущественно В12) и белковыми веществами. Слепая кишка зайцеобразных представляет собой бродильный чан для переработки клетчатки и насыщена симбиотическими микроорганизмами. В 1 г мягкого кала насчитывается до 10 млрд бактерий. Попадая вместе с фекалиями в желудок кролика, микроорганизмы полностью погибают под влиянием кислоты и перевариваются в желудке и длинном тонком кишечнике. Таким образом, у исключительно растительноядных зайцеобразных копрофагия – это способ получения незаменимых аминокислот.
Менее обязательны, но чрезвычайно существенны мутуалистические отношения между сибирской кедровой сосной и гнездящимися в кедровниках птицами – кедровкой, поползнем и кукшей. Эти птицы, питаясь семенами сосны, обладают инстинктами запасания кормов. Они прячут мелкие порции «орешков» под слой мха и лесного опада. Значительную часть запасов птицы не находят, и семена прорастают. Деятельность этих птиц способствует, таким образом, самовозобновлению кедровников, так как семена не могут прорастать на толстом слое лесной подстилки, преграждающей им доступ к почве.
Взаимовыгодны отношения растений, имеющих сочные плоды, и птиц, питающихся этими плодами и распространяющих семена, которые обычно не поддаются перевариванию. Мутуалистические отношения с муравьями складываются у многих растений: известно около 3000 видов, обладающих приспособлениями для привлечения муравьев. Типичный пример – цекропия, дерево, растущее в бассейне Амазонки. Муравьи родов Azteca и Cramatogaster заселяют пустоты в членистом стволе цекропии и питаются специальными округлыми образованиями диаметром около 1 мм – «мюллеровыми тельцами», которые растение продуцирует на вздутиях, расположенных на внешней стороне влагалища листа. Муравьи-сожители бдительно охраняют листья от вредителей, особенно от муравьев-листорезов рода Atta.
Чем разнообразнее и прочнее связи, поддерживающие совместное обитание видов, тем устойчивее их сожительство. Сообщества, имеющие длительную историю развития, поэтому прочнее, чем те, которые возникают после резких нарушений природной обстановки или создаются искусственно (поля, сады, огороды, оранжереи, теплицы, аквариумы и т. п.).
7.3.4. Нейтрализм, аменсализм
Нейтрализм – это такая форма биотических отношений, при которой сожительство двух видов на одной территории не влечет для них ни положительных, ни отрицательных последствий. При нейтрализме виды не связаны друг с другом непосредственно, но зависят от состояния сообщества в целом. Например, белки и лоси, обитая в одном лесу, практически не контактируют друг с другом. Однако угнетение леса длительной засухой либо оголение его при массовом размножении вредителей сказывается на каждом из этих видов, хотя и в неодинаковой степени. Отношения типа нейтрализма особенно развиты в насыщенных видами сообществах, включающих разных по экологии сочленов.
При аменсализме для одного из двух взаимодействующих видов последствия совместного обитания отрицательны, тогда как другой не получает от них ни вреда, ни пользы. Такая форма взаимодействия чаще встречается у растений. Например, светолюбивые травянистые виды, растущие под елью, испытывают угнетение в результате сильного затенения ее кроной, тогда как для самого дерева их соседство может быть безразличным.
Взаимосвязи этого типа также ведут к регуляции численности организмов, влияют на распределение и взаимный подбор видов.
7.3.5. Конкуренция
Конкуренция – это взаимоотношения видов со сходными экологическими требованиями существующих за счет общих ресурсов, имеющихся в недостатке. Когда такие виды обитают совместно, каждый из них находится в невыгодном положении, так как присутствие другого уменьшает возможности в овладении пищей, убежищами и прочими средствами к существованию, которыми располагает местообитание. Конкуренция – единственная форма экологических отношений, отрицательно сказывающаяся на обоих взаимодействующих партнерах.
Формы конкурентного взаимодействия могут быть самыми различными: от прямой физической борьбы до мирного совместного существования. Тем не менее если два вида с одинаковыми экологическими потребностями оказываются в одном сообществе, рано или поздно один конкурент вытесняет другого. Это одно из наиболее общих экологических правил, которое получило название закона конкурентного исключения и было сформулировано Г. Ф. Гаузе.
В упрощенной форме оно звучит как «два конкурирующих вида вместе не уживаются».
Несовместимость конкурирующих видов еще раньше была подчеркнута Ч. Дарвином, который считал конкуренцию одной из важнейших составных частей борьбы за существование, играющей большую роль в эволюции видов.
В опытах Г. Ф. Гаузе с культурами туфелек Paramecium aurelia и P. caudatum каждый из видов, помещенных отдельно в пробирки с сенным настоем, успешно размножался, достигая определенного уровня численности. Если же оба вида со сходным характером питания помещали совместно, то первое время наблюдался рост численности каждого из них, но затем количество P. caudatum постепенно сокращалось, и они исчезали из настоя, тогда как количество P. aurelia оставалось постоянным (рис. 86).
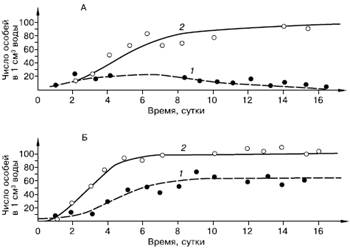
Рис. 86. Рост численности инфузорий Paramaecium caudatum (1) и P. aurelia (2) (по Г. Ф. Гаузе из Ф. Дре, 1976): А – в смешанной культуре; Б – в раздельных культурах
Победителем в конкурентной борьбе оказывается, как правило, тот вид, который в данной экологической обстановке имеет хотя бы небольшие преимущества перед другим, т. е. больше приспособлен к условиям окружающей среды, поскольку даже близкие виды никогда не совпадают по всему экологическому спектру. Так, в опытах Т. Паркас лабораторными культурами мучных хрущаков выявлено, что результат конкуренции может определяться тем, при какой температуре и влажности протекает опыт. В многочисленных стаканчиках с мукой, в которые помещали по нескольку экземпляров жуков двух видов (Tribolium confusum и Т. castaneum) и в которых они размножались, через некоторое время оставался только один из видов. При высокой температуре и влажности муки это был Т. castaneum, при более низкой температуре и умеренной влажности – Т. confusum. Однако при средних значениях факторов «победа» того или иного вида явно носила случайный характер, и предсказать исход конкуренции было трудно.
Причины вытеснения одного вида другим могут быть различны. Поскольку экологические спектры даже близких видов никогда не совпадают полностью, при общем сходстве требований к среде виды все же чем-либо отличаются друг от друга. Даже если такие виды мирно уживаются вместе, но интенсивность размножения одного чуть больше, чем другого, то постепенное исчезновение из сообщества второго вида лишь дело времени, так как с каждым поколением все больше и больше ресурсов оказывается захваченным более конкурентоспособным партнером. Часто, однако, конкуренты активно воздействуют друг на друга.
У растений подавление конкурентов происходит в результате перехвата минеральных питательных веществ и почвенной влаги корневой системой и солнечного света – листовым аппаратом, а также в результате выделения токсичных соединений. Например, в смешанных посевах двух видов клевера Trifolium repens раньше образует полог листьев, но затем его затеняет Т. fragiferum, у которого более длинные черешки. При совместном выращивании рясок Lemna gibba и Spirodela polyrrhiza численность второго вида сначала нарастает, а затем падает, хотя в чистых культурах скорость роста этого вида выше, чем первого. Преимущества L. gibba в данном случае в том, что в условиях загущения у него развивается аэренхима, помогающая держаться на поверхности воды. S. polyrrhiza, у которой нет аэренхимы, оттесняется вниз и затеняется конкурентом.
Химические взаимодействия растений через продукты их обмена веществ получили название аллелопатии. Подобные способы влияния друг на друга свойственны и животным. В приведенных выше опытах Г. Ф. Гаузе и Т. Парка подавление конкурентов происходило в основном в результате накопления в среде токсичных продуктов обмена, к которым один из видов более чувствителен, чем другой. Высшие растения с низкой потребностью в азоте, первыми появляющиеся на залежных почвах, корневыми выделениями подавляют образование клубеньков у бобовых и деятельность свободноживущих азотфиксирующих бактерий. Предотвращая тем самым обогащение почвы азотом, они получают преимущества в конкуренции с растениями, нуждающимися в большом его количестве в почве. Рогоз в зарастающих водоемах аллелопатически активен по отношению к другим водным растениям, что позволяет ему, избегая конкурентов, расти практически в чистых зарослях.
У животных могут встречаться случаи прямого нападения одного вида на другой в конкурентной борьбе. Например, личинки яйцеедов Diachasoma tryoni и Opius humilis, оказавшиеся в одном яйце хозяина, вступают друг с другом в схватку и убивают соперника, прежде чем приступить к питанию.
Возможность конкурентного вытеснения одного вида другим является результатом экологической индивидуальности видов. В неизменных условиях они будут иметь разную конкурентоспособность, так как обязательно отличаются друг от друга по толерантности к каким-либо факторам. В природе, однако, в большинстве случаев среда изменчива и в пространстве, и во времени, и это дает возможность сосуществования многих конкурентов. Например, если погодные условия более или менее регулярно меняются в пользу то одного, то другого вида, начинающиеся процессы вытеснения ими друг друга не доходят до конца и меняют знак на противоположный. Так, во влажные годы в нижнем ярусе леса могут разрастаться мхи, а в сухие их теснит покров осоки волосистой или других трав. Эти виды уживаются также в одном фитоценозе, занимая разные по условиям увлажнения участки леса. Кроме того, виды, конкурирующие не за один, а за несколько ресурсов, часто имеют разные пороги ограничивающих факторов, что также мешает завершиться процессам конкурентного исключения. Так, американский эколог Д. Тилман, культивируя совместно два вида диатомовых водорослей, выяснил, что они не вытесняют друг друга, потому что имеют разную чувствительность к недостатку азота и кремния. Вид, способный в размножении опередить другой при низком содержании азота, не может этого достичь из-за нехватки для него кремния, тогда как его конкуренту, наоборот, достаточно кремния, но мало азота.
Конкурирующие виды могут уживаться в сообществе и в том случае, если повышение численности более сильного конкурента не допускается хищником. В этом случае деятельность хищника приводит к повышению видового разнообразия сообщества. В одном из опытов со дна прибрежного участка моря, где обитало 8 видов сидячих беспозвоночных – мидий, морских желудей, морских уточек, хитонов, – удалили хищника, морскую звезду, питавшуюся в основном мидиями. Через некоторое время мидии заняли весь участок дна, вытеснив все остальные виды.
Таким образом, биоценозы содержат в каждой группе организмов значительное число потенциальных или частичных конкурентов, состоящих в динамических отношениях друг с другом. Вид может не иметь также сильных соперников, но испытывать небольшое влияние со стороны каждого из многих других, частично использующих его ресурсы. В этом случае говорят о «диффузной» конкуренции, исход которой также зависит от многих обстоятельств и может закончиться вытеснением данного вида из биоценоза.
Конкуренция, следовательно, имеет двоякое значение в биоценозах. Она является фактором, в значительной мере определяющим видовой состав сообществ, поскольку интенсивно конкурирующие виды вместе не уживаются. С другой стороны, частичная или потенциальная конкуренция позволяет видам быстро захватывать дополнительные ресурсы, освобождающиеся при ослаблении деятельности соседей, и замещать их в биоценотических связях, что сохраняет и стабилизирует биоценоз в целом.
Как и в случае любых других форм биотических связей, конкуренцию часто нелегко отделить от других типов отношений. В этом плане показательны особенности поведения экологически сходных видов муравьев.
Крупные луговые муравьи Formica pratensis строят насыпные гнезда и охраняют территорию вокруг них. У более мелких F. cunicularia гнезда небольшие, в виде земляных холмиков. Они часто селятся на периферии гнездовой территории луговых муравьев и охотятся на их кормовых участках.
При экспериментальной изоляции гнезд лугового муравья эффективность охоты F. cunicularia возрастает в 2–3 раза. Муравьи приносят более крупных насекомых, которые обычно являются добычей F. pratensis. Если изолировать гнезда F. cunicularia, добыча луговых муравьев не увеличивается, как следовало бы ожидать, а сокращается вдвое. Оказалось, что более подвижные и активные фуражиры F. cunicularia служат стимуляторами поисковой активности луговых муравьев, своеобразными разведчиками белковой пищи. Интенсивность движения фуражиров лугового муравья по дорогам в тех секторах, где есть гнезда F. cunicularia, в 2 раза выше, чем там, где их нет. Таким образом, перекрывание охотничьей территории и пищевых спектров позволяет рассматривать F. cunicularia как конкурента луговых муравьев, но повышение эффективности охоты F. pratensis свидетельствует о пользе пребывания F. cunicularia на их территории.
Все перечисленные типы биоценотических связей, выделяемые по критерию пользы или вреда взаимных контактов для отдельных партнеров, характерны не только для межвидовых, но и для внутривидовых отношений. Однако проявления их внутри вида либо развиты в иной степени, чем между разными видами, либо имеют некоторую специфику. Так, например, отношения типа хищник – жертва, паразит – хозяин у представителей одного и того же вида встречаются в природе сравнительно редко. Каннибализм, т. е. поедание себе подобных, наиболее развит у хищных рыб – щук, окуней, корюшки, трески, наваги и др. В условиях обостренной конкуренции за пищу или воду каннибализм проявляется иногда и у нехищных животных. Например, личинки майского жука, помещенные в сухую почву, могут поедать друг друга. Паразитирование на себе подобных встречается еще реже и характеризует в основном отношения полов. У сидячих боннелий, относящихся к родственной кольчецам группе эхиурид, карликовый самец паразитирует на относительно крупной, самостоятельно питающейся самке. Подобные отношения характерны и для некоторых глубоководных рыб-удильщиков (рис. 87). Самки носят на себе значительно более мелких самцов, которые прирастают ртом к их телу и питаются как паразиты. Такой внутривидовой паразитизм имеет приспособительное значение: наличие «карманных» самцов снимает необходимость затраты энергии на встречу полов, а при малой величине самца снижает конкуренцию из-за пищи в условиях общего недостатка кормов на больших глубинах.
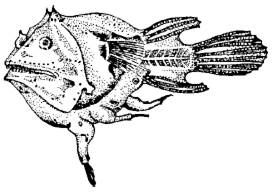
Рис. 87. Самка глубоководного удильщика с тремя приросшими к ней самцами
Мутуалистические и конкурентные отношения представляют собой основную сущность внутривидовых связей. Изучение роли этих взаимоотношений в пределах вида, многообразия и специфики их форм является предметом специального раздела синэкологии – экологии популяций.
Как видно из приведенных примеров, формальная классификация типов биотических связей не может полностью отразить все их разнообразие и сложность в живой природе, но все же позволяет ориентироваться в главных типах взаимодействия организмов. Другие классификации обращают внимание на иные аспекты биотических отношений, используя другие подходы.
В. Н. Беклемишев подразделял отношения между видами в сообществе на прямые и косвенные. Прямые связи возникают при непосредственном контакте организмов. Косвенные связи представляют собой влияние видов друг на друга через среду обитания или путем воздействия на третьи виды.
По классификации В. Н. Беклемишева, прямые и косвенные межвидовые отношения по тому значению, которое они могут иметь в биоценозе, подразделяются на четыре типа: трофические, топические, форические, фабрические.
7.3.6. Трофические связи
Трофические связи возникают, когда один вид питается другим – либо живыми особями, либо их мертвыми остатками, либо продуктами жизнедеятельности. И стрекозы, ловящие на лету других насекомых, и жуки-навозники, питающиеся пометом крупных копытных, и пчелы, собирающие нектар растений, вступают в прямую трофическую связь с видами, предоставляющими им пищу. В случае конкуренции двух видов из-за объектов питания между ними возникает косвенная трофическая связь, так как деятельность одного отражается на снабжении кормом другого. Любое воздействие одного вида на поедаемость другого или доступность для него пищи следует расценивать как косвенную трофическую связь между ними. Например, гусеницы бабочек-монашенок, объедая хвою сосен, облегчают короедам доступ к ослабленным деревьям.
Трофические связи являются главными в сообществах. Именно они объединяют живущие вместе виды, поскольку каждый из них может обитать лишь там, где имеются необходимые ему пищевые ресурсы. Любой вид не только приспособлен к определенным источникам питания, но и сам служит пищевым ресурсом для других. Пищевые взаимосвязи создают в природе трофическую сеть, распространяющуюся в конечном счете на все виды в биосфере. Образ этой трофической сети можно воссоздать, поставив в центр любой вид и соединив его стрелками со всеми другими, которые находятся с ним в прямых или косвенных пищевых отношениях (рис. 88), и затем продолжать эту процедуру для каждого вовлеченного в схему вида. В результате будет охвачена вся живая природа, от китов до бактерий. Как показали исследования академика А. М. Уголева, существует «чрезвычайное единообразие свойств ассимиляционных систем на молекулярном и надмолекулярном уровне у всех организмов биосферы», позволяющее им получать энергетические ресурсы друг от друга. Он утверждает, что за бесконечным разнообразием типов питания стоят общие фундаментальные процессы образующих единую систему трофических взаимодействий планетарного масштаба.
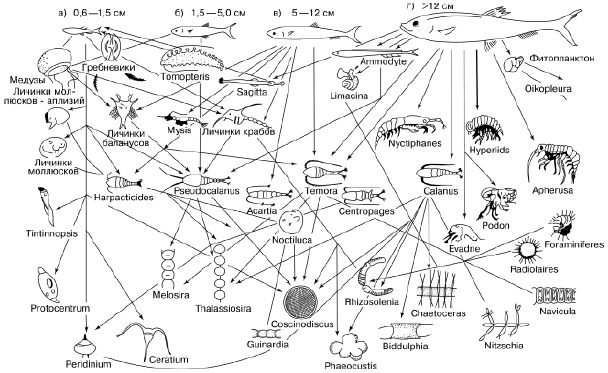
Рис. 88. Пищевые связи сельди – часть трофической сети океана
Любой биоценоз пронизан пищевыми связями и представляет собой более или менее локализованный в пространстве участок общей трофической сети, связывающей все живое на Земле.
7.3.7. Топические связи
Топические связи характеризуют любое, физическое или химическое, изменение условий обитания одного вида в результате жизнедеятельности другого. Эти связи крайне разнообразны. Они заключаются в создании одним видом среды для другого (например, внутренний паразитизм или норовый комменсализм), в формировании субстрата, на котором поселяются или, наоборот, избегают селиться представители других видов, во влиянии на движение воды, воздуха, изменение температуры, освещенности окружающего пространства, в насыщении среды продуктами выделения и т. п. Морские желуди, поселяющиеся на коже китов, личинки мух, обитающие в лепешках коровьего навоза, лишайники на стволах деревьев связаны прямой топической связью с теми организмами, которые предоставляют им субстрат или среду обитания. Особенно большая роль в создании или изменении среды для других организмов принадлежит растениям. Растительность из-за особенностей энергообмена является мощным фактором перераспределения тепла у поверхности Земли и создания мезо– и микроклимата. Под пологом леса подлесок, напочвенный покров, а также все животное население находятся в условиях более выровненных температур, более высокой влажности воздуха и т. д. Травянистая растительность, хотя и в меньшей степени, также изменяет режим окружающего пространства. В степях около дерновин ковыля перистого и овсяницы Беккера температура поверхности почвы с теневой стороны может быть на 8-12 °C ниже, чем на незаросших участках. Здесь концентрируются многие мелкие насекомые. В результате отрицательных или положительных топических взаимоотношений одни виды определяют или исключают возможность существования в биоценозе других видов.
На основе топических связей в биоценозе формируются консорции (сочетания) – группы разнородных организмов, поселяющихся на теле или в теле особи какого-либо определенного вида – центрального члена консорции. В большинстве случаев члены одной консорции связаны также разнообразными трофическими отношениями. Консорции формируются фактически вокруг представителей любого вида, обладающего средообразующим воздействием на других. По свидетельству В. Н. Беклемишева, «сосна с ее микоризными грибками, с эпифитными мхами и лишайниками на стволе и ветвях, со всем множеством населяющих ее членистоногих – это сложнейший консорций, это – целый мир. Полевка с ее эктопаразитами, с гельминтами, с простейшими, бактериями, населяющими ее внутренние органы, опять есть целый консорций».
Отдельные консорции могут быть разной степени сложности. Наибольшим числом консортивных связей отличаются те растения, которым принадлежит основная роль в создании внутренней среды биоценоза. Так как каждый член крупной консорции может быть, в свою очередь, центром более мелкого объединения, можно выделить консорции первого, второго и даже третьего порядка. Таким образом, биоценоз – это система связанных между собой консорций, возникающих на основе теснейших топических и трофических отношений между видами. Консортивные связи, в основе которых лежат топические отношения, формируют своего рода блочную структуру биоценоза.
Топические и трофические связи имеют наибольшее значение в биоценозе, составляют основу его существования. Именно эти типы отношений удерживают друг возле друга организмы разных видов, объединяя их в достаточно стабильные сообщества разных масштабов.
7.3.8. Форические связи
Форические связи – это участие одного вида в распространении другого. В роли транспортировщиков выступают животные. Перенос животными семян, спор, пыльцы растений называют зоохорией, перенос других, более мелких животных – форезией (от лат. форас– наружу, вон). Перенос осуществляется обычно с помощью специальных и разнообразных приспособлений. Животные могут захватывать семена растений двумя способами: пассивным и активным. Пассивный захват происходит при случайном соприкосновении тела животного с растением, семена или соплодия которого обладают специальными зацепками, крючками, выростами (череда, лопух). Распространителями их обычно служат млекопитающие, которые на шерсти переносят такие плоды иногда на довольно значительные расстояния. Активный способ захвата – поедание плодов и ягод. Не поддающиеся перевариванию семена животные выделяют вместе с пометом. В переносе грибных спор большую роль играют насекомые. По-видимому, плодовые тела грибов возникли как образования, привлекающие насекомых-расселителей.
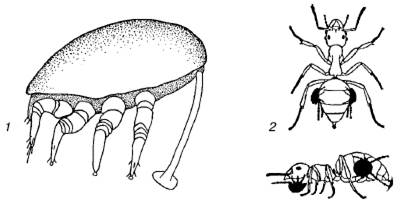
Рис. 89. Форезия клещей на насекомых:
1– дейтонимфа уроподового клеща прикрепляется к жуку стебельком из затвердевшей секреторной жидкости;
2– форезия клещей на муравьях
Форезия животных распространена преимущественно среди мелких членистоногих, особенно у разнообразных групп клещей (рис. 89). Она представляет собой один из способов пассивного расселения и свойственна видам, для которых перенос из одного биотопа в другой жизненно необходим для сохранения или процветания. Например, многие летающие насекомые – посетители скоплений быстро разлагающихся растительных остатков (трупов животных, помета копытных, куч гниющих растений и т. п.) несут на себе гамазовых, уроподовых или тироглифоидных клещей, переселяющихся таким образом от одного скопления пищевых материалов к другому. Собственные расселительные возможности не позволяют этим видам преодолевать значительные для них расстояния. Жуки-навозники иногда ползают с поднятыми надкрыльями, которые не в состоянии сложить из-за густо усеявших тело клещей. Посредством форезии на насекомых распространяются некоторые виды нематод (рис. 90). Ноги навозных мух часто имеют вид ламповых щеток из-за обилия прикрепившихся к ним нематод рабдитид. Среди крупных животных форезия почти не встречается.

Рис. 90. Расселение личинок нематод на жуках:
1– личинки в ожидании расселителя;
2– личинки, прикрепившиеся под надкрыльями жука
7.3.9. Фабрические связи
Фабрические связи – это такой тип биоценотических отношений, в которые вступает вид, использующий для своих сооружений (фабрикаций) продукты выделения, либо мертвые остатки, либо даже живых особей другого вида. Так, птицы употребляют для постройки гнезд ветви деревьев, шерсть млекопитающих, траву, листья, пух и перья других видов птиц и т. п. Личинки ручейников строят домики из кусочков веток, коры или листьев растений, из раковин мелких видов катушек, захватывая даже раковинки с живыми моллюсками. Пчела-мегахила помещает яйца и запасы в стаканчики, сооружаемые из мягких листьев различных кустарников (шиповника, сирени, акации и т. п.).
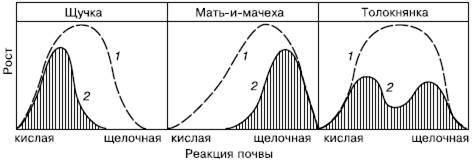
Рис. 91. Схема влияния рН на рост различных растений при выращивании в одновидовых посевах и в условиях конкуренции:
1– кривые физиологического оптимума;
2– синэкологического оптимума (по В. Лархеру, 1978)
Из-за сложности межвидовых отношений каждый конкретный вид может преуспевать далеко не везде, где складываются подходящие для него условия физической среды. Различают физиологический и синэкологический оптимумы вида. Физиологический оптимум – это благоприятное для вида сочетание всех абиотических факторов, при котором возможны наиболее быстрые темпы роста и размножения. Синэкологический оптимум – это такое биотическое окружение, при котором вид испытывает наименьшее давление со стороны врагов и конкурентов, что позволяет ему успешно размножаться. Синэкологический и физиологический оптимумы далеко не всегда совпадают (рис. 91). Если в подходящем биотопе экологическая ниша занята более сильным конкурентом или чересчур велико влияние хищников и паразитов, вид в нем не приживается. Пример несовпадения физиологического и синэкологического оптимумов – массовое размножение вредителя зерновых культур, гессенского комарика, после особенно суровых зим, которые, казалось бы, должны неблагоприятно сказываться на численности этого насекомого. В нормальные по условиям годы гессенского комарика сильно истребляют несколько видов его естественных врагов – паразитических перепончатокрылых наездников. В суровые зимы из-за очень слабой морозоустойчивости враги гессенского комарика вымерзают почти полностью. Это дает возможность вредителю быстро восстановить собственную численность, сокращенную морозами, и беспрепятственно размножаться в угрожающем для урожая количестве.
Экологическая ниша
Положение вида, которое он занимает в общей системе биоценоза, комплекс его биоценотических связей и требований к абиотическим факторам среды называют экологической нишей вида.
Концепция экологической ниши оказалась очень плодотворной для понимания законов совместной жизни видов. Над ее развитием работали многие экологи: Дж. Гриннелл, Ч. Элтон, Г. Хатчинсон, Ю. Одум и др.
Понятие «экологическая ниша» следует отличать от понятия «местообитание». В последнем случае подразумевается та часть пространства, которая заселена видом и которая обладает необходимыми абиотическими условиями для его существования. Экологическая ниша вида зависит не только от абиотических условий среды, но и в не меньшей мере от его биоценотического окружения. Характер занимаемой экологической ниши определяется как экологическими возможностями вида, так и тем, насколько эти возможности могут быть реализованы в конкретных биоценозах. Это характеристика того образа жизни, который вид может вести в данном сообществе.
Г. Хатчинсон выдвинул понятия фундаментальной и реализованной экологической ниши. Под фундаментальной понимается весь набор условий, при которых вид может успешно существовать и размножаться. В природных биоценозах, однако, виды осваивают далеко не все пригодные для них ресурсы вследствие, прежде всего, конкурентных отношений. Реализованная экологическая ниша – это положение вида в конкретном сообществе, где его ограничивают сложные биоценотические отношения. Иными словами, фундаментальная экологическая ниша характеризует потенциальные возможности вида, а реализованная – ту их часть, которая может осуществиться в данных условиях, при данной доступности ресурса. Таким образом, реализованная ниша всегда меньше, чем фундаментальная.
В экологии широко обсуждается вопрос о том, сколько экологических ниш может вместить биоценоз и сколько видов какой-либо конкретной группы, имеющих близкие требования к среде, могут ужиться вместе.
Специализация вида по питанию, использованию пространства, времени активности и другим условиям характеризуется как сужение его экологической ниши, обратные процессы – как ее расширение. На расширение или сужение экологической ниши вида в сообществе большое влияние оказывают конкуренты. Правило конкурентного исключения, сформулированное Г. Ф. Гаузе для близких по экологии видов, может быть выражено таким образом, что два вида не уживаются в одной экологической нише.
Эксперименты и наблюдения в природе показывают, что во всех случаях, когда виды не могут избежать конкуренции за основные ресурсы, более слабые конкуренты постепенно вытесняются из сообщества. Однако в биоценозах возникает много возможностей хотя бы частичного разграничения экологических ниш близких по экологии видов.
Выход из конкуренции достигается благодаря расхождению требований к среде, изменению образа жизни, что, другими словами, является разграничением экологических ниш видов. В этом случае они приобретают способность сосуществовать в одном биоценозе. Каждый из живущих вместе видов в отсутствие конкурента способен на более полное использование ресурсов. Это явление легко наблюдать в природе. Так, травянистые растения ельника способны довольствоваться небольшим количеством почвенного азота, которое остается от перехвата его корнями деревьев. Однако если на ограниченной площадке обрубить корни этих елей, условия азотного питания трав улучшаются и они бурно идут в рост, принимая густо-зеленую окраску. Улучшение условий жизни и увеличение численности какого-либо вида в результате удаления из биоценоза другого, близкого по экологическим требованиям, называется конкурентным высвобождением.
Разделение совместно живущими видами экологических ниш с частичным их перекрыванием – один из механизмов устойчивости природных биоценозов. Если какой-либо из видов резко снижает свою численность или выпадает из состава сообщества, его роль берут на себя другие. Чем больше видов в составе биоценоза, тем ниже численность каждого из них, тем сильнее выражена их экологическая специализация. В этом случае говорят о «более плотной упаковке экологических ниш в биоценозе».
У близкородственных видов, живущих вместе, обычно наблюдаются очень тонкие разграничения экологических ниш. Так, пасущиеся в африканских саваннах копытные по-разному используют пастбищный корм: зебры обрывают в основном верхушки трав, антилопы гну кормятся тем, что оставляют им зебры, выбирая при этом определенные виды растений, газели выщипывают самые низкие травы, а антилопы топи довольствуются высокими сухими стеблями, оставшимися после других травоядных. Такое же «разделение труда» в южноевропейских степях осуществляли когда-то дикие лошади, сурки и суслики (рис. 92).
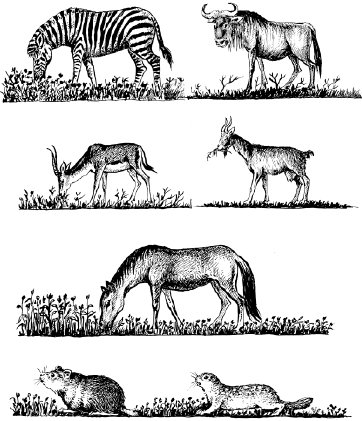
Рис. 92. Разные виды травоядных поедают траву на разной высоте в африканских саваннах (верхние ряды) и в степях Евразии (нижние ряды) (по Ф. Р. Фуэнте, 1972; Б. Д. Абатурову, Г. В. Кузнецову, 1973)
В наших зимних лесах насекомоядные птицы, кормящиеся на деревьях, также избегают конкуренции друг с другом за счет разного характера поиска. Например, поползни и пищухи собирают пищу на стволах. При этом поползни стремительно обследуют дерево, быстро схватывая попадающихся на глаза насекомых или семена, оказавшиеся в крупных трещинах коры, тогда как мелкие пищухи тщательно обшаривают на поверхности ствола малейшие щелки, в которые проникает их тонкий шиловидный клюв. Зимой в смешанных стайках большие синицы ведут широкий поиск на деревьях, в кустах, на пнях, а часто и на снегу; синицы-гаички обследуют преимущественно крупные ветви; длиннохвостые синицы ищут корм на концах ветвей; мелкие корольки тщательно обшаривают верхние части крон хвойных.
Муравьи существуют в природных условиях многовидовыми ассоциациями, члены которых различаются по образу жизни. В лесах Подмосковья чаще всего обнаруживается такая ассоциация видов: доминантный вид (Formica rufa, F. aquilonia или Lasius fuliginosus) занимает несколько ярусов, в почве активен L. flavus, в подстилке леса – Myrmica rubra, напочвенный ярус осваивают L. niger и F. fusca, деревья – Camponotus herculeanus. Специализация к жизни в разных ярусах отражается в жизненной форме видов. Кроме разделения в пространстве, муравьи отличаются и по характеру добывания пищи, по времени суточной активности.
В пустынях наиболее развит комплекс муравьев, собирающих пищу на поверхности почвы (герпетобионтов). Среди них выделяются представители трех трофических групп: 1) дневные зоонекрофаги – активны в самое жаркое время, питаются трупами насекомых и активными днем мелкими живыми насекомыми; 2) ночные зоофаги – охотятся на малоподвижных насекомых с мягкими покровами, которые появляются на поверхности только ночью, и на линяющих членистоногих; 3) карпофаги (дневные и ночные) – поедают семена растений.
Совместно могут обитать по нескольку видов из одной трофической группы. Механизмы выхода из конкуренции и разграничения экологических ниш при этом следующие.
1. Размерная дифференциация (рис. 93). Например, средние веса рабочих особей трех наиболее обычных в песках Кызылкумов дневных зоонекрофагов относятся как 1:8:120. Примерно такое же соотношение весов у некрупной кошки, рыси и тигра.
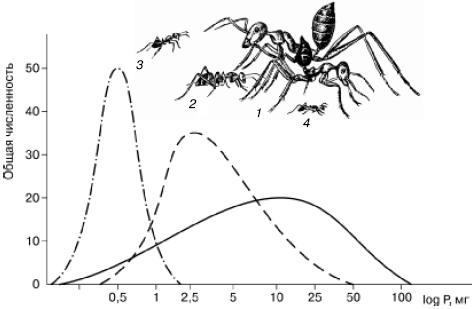
Рис. 93. Сравнительные размеры четырех видов муравьев из группы дневных зоонекрофагов в песчаной пустыне Центральных Каракумов и распределение добычи трех видов по весовым классам (по Г. М. Длусскому, 1981): 1– средний и крупный рабочие Cataglyphis setipes; 2 – С. pallida; 3 – Acantholepis semenovi; 4 – Plagiolepis pallescens
2. Поведенческие различия заключаются в разной стратегии фуражировки. Муравьи, которые создают дороги и используют мобилизацию носильщиков для переноса в гнездо обнаруженной пищи, питаются преимущественно семенами растений, образующих куртины. Муравьи, фуражиры которых работают как одиночные сборщики, собирают в основном семена растений, распределенных дисперсно.
3. Пространственная дифференциация. В пределах одного яруса сбор пищи разными видами может быть приурочен к разным участкам, например на открытых местах или под кустиками полыни, на песчаных или глинистых площадках и т. д.
4. Различия во времени активности относятся преимущественно ко времени суток, но у некоторых видов отмечены несовпадения активности и по сезонам года (преимущественно весенняя или осенняя активность).
Экологические ниши видов изменчивы в пространстве и во времени. Они могут быть резко разграничены в индивидуальном развитии в зависимости от стадии онтогенеза, как, например, у гусениц и имаго чешуекрылых, личинок и жуков майского хруща, головастиков и взрослых лягушек. В этом случае меняется и среда обитания, и все биоценотическое окружение. У других видов экологические ниши, занимаемые молодыми и взрослыми формами, более близки, но тем не менее между ними всегда имеются различия. Так, взрослые окуни и их мальки, живущие в одном и том же озере, используют для своего существования разные энергетические источники и входят в разные цепи питания. Мальки живут за счет мелкого планктона, взрослые – типичные хищники.
Ослабление межвидовой конкуренции приводит к расширению экологической ниши вида. На океанических островах с бедной фауной ряд птиц по сравнению с их сородичами на материке заселяет более разнообразные местообитания и расширяет спектр кормов, так как не сталкивается при этом с конкурирующими видами. У островных обитателей отмечается даже повышенная изменчивость формы клюва как показатель расширения характера кормовых связей.
Если межвидовая конкуренция сужает экологическую нишу вида, не давая проявиться всем его потенциям, то внутривидовая конкуренция, наоборот, способствует расширению экологических ниш. При возросшей численности вида начинается использование дополнительных кормов, освоение новых местообитаний, появление новых биоценотических связей.
Экологические ниши растений, на первый взгляд, менее разнообразны, чем животных. Они четко очерчены у видов, различающихся по питанию: сапрофитов (подъельник, гнездовка), паразитов (заразиха, повилика, раффлезия), полупаразитов (погремок, марьянник, омела), насекомоядных растений (росянка, пузырчатка, непентес). Но и зеленые фотоавтотрофные растения занимают разные экологические ниши в фитоценозах. Множественность экологических ниш определяется разным использованием растениями среды, размещением органов в воздухе и почве, ритмами сезонного развития, длительностью периода вегетации, сроками цветения и особенностями плодоношения, взаимосвязями с элементами абиотической среды и всеми компонентами биоценоза.
В водоемах растения, полностью погруженные в воду (элодея, роголистник, уруть), оказываются в иных условиях температуры, освещенности, газового режима, чем плавающие на поверхности (телорез, водокрас, ряска) или укореняющиеся на дне и выносящие листья на поверхность (кувшинка, кубышка, виктория). Различаются они и взаимосвязями со средой. Эпифиты тропических лесов занимают сходные, но все же не идентичные ниши, так как относятся к разным экологическим группам по отношению к свету и воде (гелиофиты и сциофиты, гигрофиты, мезофиты и ксерофиты). Разные эпифитные орхидеи имеют узкоспециализированных опылителей.
В зрелом широколиственном лесу деревья первого яруса – дуб обыкновенный, вяз гладкий, клен платановидный, липа сердцелистная, ясень обыкновенный имеют сходные жизненные формы. Древесный полог, образованный их кронами, оказывается в одном горизонте, в сходных условиях среды. Но внимательный анализ показывает, что они по-разному участвуют в жизни сообщества и, следовательно, занимают разные экологические ниши. Эти деревья различаются по степени светолюбия и теневыносливости, срокам цветения и плодоношения, способам опыления и распространения плодов, составу консортов и проч. Дуб, вяз и ясень – анемофильные растения, но насыщение среды их пыльцой происходит в разные сроки. Клен и липа – энтомофилы, хорошие медоносы, но цветут в разное время. У дуба – зоохория, у остальных широколиственных деревьев – анемохория. Состав консортов у всех разный.
Если в широколиственном лесу кроны деревьев находятся в одном горизонте, то активные корневые окончания располагаются на разной глубине. Корни дуба проникают наиболее глубоко, выше располагаются корни клена и еще более поверхностно – ясеня. Опад разных видов деревьев утилизируется с разной скоростью. Листья липы, клена, вяза, ясеня к весне почти полностью разлагаются, а листья дуба и весной еще образуют рыхлую лесную подстилку.
В соответствии с представлениями Л. Г. Раменского об экологической индивидуальности видов и с учетом того, что виды растений в сообществе по-разному участвуют в освоении и преобразовании среды и трансформации энергии, можно считать, что в сложившихся фитоценозах каждый вид растения имеет свою экологическую нишу.
В онтогенезе растения, как и многие животные, меняют экологическую нишу. С возрастом они более интенсивно используют и преобразуют среду. Переход растения в генеративный период заметно расширяет круг консортов, меняет размер и напряженность фитогенного поля. Средообразующая роль стареющих, сенильных растений убывает. Они теряют многих консортов, но увеличивается роль связанных с ними деструкторов. Продукционные процессы ослабляются.
У растений имеет место перекрывание экологических ниш. Оно усиливается в отдельные периоды при ограничении ресурсов среды, но поскольку виды используют ресурсы индивидуально, избирательно и с разной интенсивностью, конкуренция в устойчивых фитоценозах ослабляется.
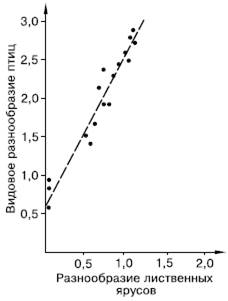
Рис. 94. Корреляция между разнообразием лиственных ярусов и видовым разнообразием птиц (индексы Шеннона по Макартуру из Э. Пианка, 1981)
На богатство экологических ниш в биоценозе оказывают влияние две группы причин. Первая – условия среды, предоставляемые биотопом. Чем мозаичнее и разнообразнее биотоп, тем больше видов могут размежевать в нем свои экологические ниши. Другой источник разнообразия ниш – сами виды, являющиеся ресурсом и создающие среду для других. Американский исследователь Р. Макартур, изучая ярусное разнообразие растительности по горному склону и число видов гнездящихся птиц, продемонстрировал прямую зависимость между этими показателями, положив начало многим исследованиям, подтверждающим связь видовой емкости сообществ с их внутренней структурой (рис. 94). Любой новый вид, внедряющийся в сообщество, увеличивает число имеющихся в нем экологических ниш не только за счет своего собственного положения среди других, но и предоставляя ресурсы для паразитов и хищников, норовых и гнездовых сожителей и т. п.
Дата добавления: 2016-03-22; просмотров: 1016;