Альтернативные структуры ДНК
Рассмотренную структуру ДНК обычно считают одним из несомненных фактов молекулярной биологии. Но недавно стало очевидным, что некоторые параметры классической В-формы нужно пересмотреть и даже что ДНК может образовывать другие типы двуспиральных структур.
Проблема определения точных значений параметров двойной спирали связана с тем, что все первоначальные значения были получены при изучении дифракции рентгеновских лучей на волокнах ДНК. По этим данным установлены основные характеристики, такие, как число пар оснований на виток, расстояние между соседними парами вдоль оси спирали. Однако этим методом нельзя определить положения отдельных атомов, как это делают, используя рентгеноструктурный анализ. Поэтому модель уточняют, согласуя расчетную картину дифракции рентгеновских лучей с экспериментальными данными. Теоретически на основе одних и тех же данных всегда можно построить разные модели. Дело в том, что модель удовлетворяет усредненным данным и, следовательно, в определенных областях структуры возможны вариации. Например, до сих пор мы говорили о ДНК как о длинной, жесткой двойной спирали, но мы знаем, что в действительности она должна быть свернута и плотно уложена, для того чтобы уместиться в клетке. При этом детали ее структуры могут изменяться.
Сейчас считается, что двухцепочечная ДНК-всегда двойная спираль. Однако была предложена и другая модель, хорошо согласующаяся с данными дифракции рентгеновских лучей. Согласно этой модели, две антипараллельные цепи ДНК также соединяются путем комплементарного спаривания оснований, но лежат «бок о бок», вместо того, чтобы закручиваться в непрерывную двойную спираль.
Существование ДНК в виде двойной спирали было подтверждено экспериментами, в которых прямо измеряли число пар оснований на виток. Оказалось, что их 10,4 вместо 10,0, предсказанных классической В-моделью. Это различие вызвало необходимость несколько изменить угол вращения между соседними парами оснований вдоль спирали до 34,6°, так что отрезок спирали, в пределах которого совершается полный виток на 360°, стал несколько длиннее.
Особенно важно, что значение 10,4 является средним для ДНК как целой молекулы при определенных условиях. Изменение условий или даже последовательности отдельных оснований может привести к большему или меньшему закручиванию спиральной структуры в соответствующих участках.
Действительно, методом рентгеноструктурного анализа было показано, что молекула, состоящая из 12 пар оснований, содержит 10,1 пары оснований на виток, что обеспечивается слабым сдвигом каждой пары оснований, при котором улучшаются межплоскостные (стэкинг) взаимодействия между основаниями, по сравнению с первоначальной моделью.
Ввиду таких вариаций идея о существовании единственной структуры двуспиральной ДНК сменилась представлением, допускающим наличие семейства структур, каждая из которых имеет характерный тип, но проявляет различия по главным параметрам - n (число нуклеотидов на виток) и h (расстояние между соседними повторяющимися элементами). Вариации обусловлены изменением вращения отдельных групп вокруг связей, обладающих свободой вращения. В пределах каждого семейства структур параметры могут слабо варьировать. Например, для В-ДНК значение n может составлять от 10,0 до 10,6.
В течение долгого времени были известны три структурные формы ДНК, способные к взаимопревращениям при изменении соответствующих условий. В-форма спирали, для которой Уотсон и Крик построили свою модель, характерна для волокон ДНК при очень высокой относительной влажности (92%) и в растворах низкой ионной силы. Считают, что именно в такой форме ДНК обычно находится в живой клетке.
А-форма обнаружена в волокнах ДНК при 75% влажности и нуждается в присутствии ионов натрия, калия или цезия, несущих противоположный заряд. Основания, располагавшиеся строго перпендикулярно оси спирали в В-форме, в А-форме наклонены по отношению к оси спирали, и их число на виток больше. А-форма интересна с биологической точки зрения, так как ее конформация очевидно, близка к структуре гибридов ДНК-РНК и двуспиральных участков РНК (обсуждается далее). Причина этого заключается в том, что 2'-гидроксильная группа мешает ДНК принять В-форму.
С-форма образуется, когда ДНК находится при 66% влажности в присутствии ионов лития. У нее меньше пар оснований на виток, чем у В-ДНК.
В этих трех формах могут находиться все ДНК независимо от их нуклеотидной последовательности. Следующие формы характерны только для молекул ДНК с определенными особенностями в составе пар оснований.
D-форма и Е-форма (возможно, крайние варианты одной и той же формы) имеют наименьшее число пар на виток (8 и 7,5) и обнаружены только в определенных молекулах ДНК, не содержащих гуанина.
Z-форма представляет собой наиболее резкий контраст с классическим структурным семейством. Эта форма левоспиральная, тогда как все остальные - правоспиральные. Она имеет наибольшее число пар на виток, т.е. менее скрученная и более тонкая. Свое название форма получила из-за зигзагообразной (zigzag) линии, которую образует сахарофосфатный остов вдоль спирали. В отличие от этого остов В-формы ДНК образует плавно изгибающуюся линию, что хорошо видно на рис., где сравниваются В- и Z-формы ДНК.
Z-форма двойной спирали найдена в полимерах, образованных чередующимися пурин-пиримидиновыми последовательностями. Два исследованных полимера состояли из многократно повторенных динуклеотидных пар: poly-d (GC/CG) и poly-d (AC/TG). Буква «d» указывает, что это дезоксиформы, т.е. что это ДНК, а не РНК. Верхний и нижний ряды букв соответствуют двум цепям ДНК.
Z-форма существует только при очень высоких концентрациях соли (что объясняется необходимостью противостоять повышенному электростатическому отталкиванию между нуклеотидами, сжатыми из-за уменьшения диаметра двойной спирали Z-ДНК).
Вполне возможно, что Z-форма ДНК имеет определенное биологическое значение. Об этом свидетельствуют следующие данные: замена остатка С в полимере poly-d(GC/CG) и poly-d на 5-метилцитозин значительно повышает стабильность Z-ДНК при низкой концентрации соли. Единственное различие между 5-Ме-С и С заключается в наличии метильной группы при С-5. Эта модификация цитозина происходит in vivo в результате метилирования динуклеотидной последовательности GC/CG в некоторых участках ДНК. Отсюда вытекает, что Z-ДНК может существовать in vivo при подходящих условиях, т.е. когда последовательность соответствующего состава окажется в условиях, способствующих образованию Z-формы.
До сих пор мы рассматривали ДНК как структуру совершенно изолированную. В действительности она связана с белками, которые могут оказывать значительное влияние на возможность перехода из В-формы в Z-форму. Например, ДНК, связанная с гистонами (основные хромосомные белки эукариотического ядра), не переходит из одной формы в другую в тех условиях, когда это наблюдают у свободной ДНК. Таким образом, одним из условий, необходимых для образования Z-ДНК in vivo, по-видимому, является присутствие особых белков, стабилизирующих ее структуру.
Является ли Z-ДНК единственной левосторонней формой двойной спирали? Модели для левосторонних вариантов традиционных А-, В- и С-форм построены так, что, возможно, они и существуют. Следует помнить, что все эти формы сохраняют самую существенную в биологическом смысле особенность двойной спирали: комплементарность спаривания оснований, определяющую специфичность соединения цепей.
При построении моделей каждую форму ДНК рассматривают независимо как конструкцию, в которой находится вся молекула в определенных условиях. Но вряд ли это действительно имеет место in vivo. Большинство клеточной ДНК находится, по-видимому, в В-форме с определенными модификациями параметров спирали, варьирующими локально. И только отдельные короткие участки спирали переходят в другие формы.
А-ДНК 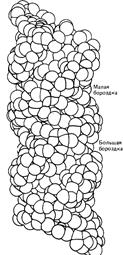 В -ДНК
В -ДНК 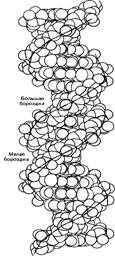 Z-ДНК
Z-ДНК 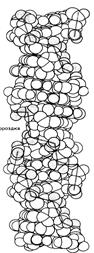
Рис. Нуклеиновые кислоты могут образовывать несколько типов двойной спирали.

А-ДНК В-ДНК Z-ДНК
Таким образом, ДНК может существовать в виде структурных семейств нескольких типов, в зависимости от концентрации ионов и нуклеотидного состава молекулы. Общие свойства этих форм суммированы в табл. 1.
| Тип спирали | Число пар оснований на виток | Угол вращения1 одной пары, градусы | Расстояние между парами оснований, А | Диаметр спирали, А |
| А | + 32,7 | 2,56 | ||
| В | + 36,0 | 3,38 | ||
| С | 9, 3 | + 38,6 | 3,32 | |
| Z | -30,0 | 3,71 |
1 Угол вращения показан знаком ( + ) в случае правосторонней спирали и знаком (—) в случае левосторонней спирали.
Репликация ДНК
Способность к репликации (удвоению) ДНК определяет ее уникальные свойства по хранению и передаче наследственной информации.
Принцип репликации ДНК состоит в том, что каждая из двух полинуклеотидных нитей молекулы ДНК служит в качестве программы (матрицы) для синтеза новой (комплементарной) нити. В результате на основе одной двухцепочечной молекулы образуются две одинаковые двухцепочечные молекулы, в каждой из которых одна цепочка является старой, а другая - новой (вновь синтезированной). Такой принцип репликации ДНК был назван полуконсервативным (рис.).
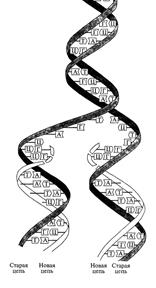
Рис. Принцип полуконсервативной репликации ДНК
В соответствии с этим принципом нуклеотидная последовательность матричной (родительской) нити считывается в направлении 3'→ 5', тогда как синтез новой (дочерней) нити идет в направлении 5' → 3'. Поскольку две комплементарные цепочки родительской молекулы ДНК являются антипараллельными, то синтез новой полинуклеотидной цепочки на каждой из них идет в противоположном направлении.
Механизм репликации ДНК является достаточно сложным и, различается в случае организмов, содержащих относительно небольшие по размерам молекулы ДНК в замкнутой (кольцевой) форме (многие вирусы и бактерии), и эукариот, клетки которых имеют молекулы огромных размеров, находящиеся в линейной (незамкнутой) форме.
У прокариотов кольцевая молекула ДНК представляет собой одну структурную единицу репликации (репликон), имеющую единственную точку начала (инициации) репликации (О-пункт, coстоящий примерно из 300 нуклеотидов), в которой начинается процесс расхождения (расплетания) двух нитей родительской молекулы и матричного синтеза комплементарных копий (реплик) дочерней ДНК.
Сайт 0-репликации (ориджин репликации, сайт инициации репликации, точка инициации репликации)
Этот процесс продолжается непрерывно по длине копируемой структуры и заканчивается в этом же репликоне образованием двух молекул «полуконсервативного» типа.
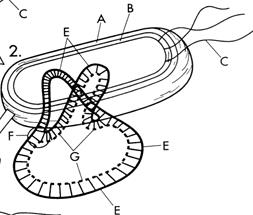
А- клеточная стенка бактерии; В –мембрана бактериальной клетки; С- жгутики; D-цитоплазма; F- энзим; Е – исходная ДНК; G- новая ДНК; О - место начала репликации.
В больших линейных молекулах ДНК эукариот имеется много точек начала репликации и соответствующих им репликонов (от нескольких сотен до десятков тысяч), т. е. такая ДНК является полирепликонной.
При рассмотрении современных представлений о механизме репликации ДНК эукариот можно условно выделить три последовательных этапа этого процесса, происходящего в репликоне, в каждом из которых принимают участие те или иные белки (ферменты).
Первый этап связан с быстрым раскручиванием двух полинуклеотидных нитей спирализованной молекулы ДНК на определенном ее участке (в границах работающего репликона) и с их разделением путем разрушения водородных связей между парами комплементарных оснований. При этом образуются два одноцепочечных фрагмента родительской молекулы, каждый из которых может выступать в роли матрицы для синтеза комплементарной (дочерней) нити. Этот этап инициируется в соответствующей точке начала репликации и обеспечивается комплексным участием нескольких различных белков. В результате их действия формируется T-образная структура, названная вилкой репликации, в которой две родительские цепочки ДНК уже отделены друг от друга (рис.). Образовавшаяся вилка репликации быстро продвигается вдоль двойной спирали родительской молекулы ДНК благодаря активности «расплетающего» фермента ДНК-хеликазы и при участии группы дестабилизирующих белков.
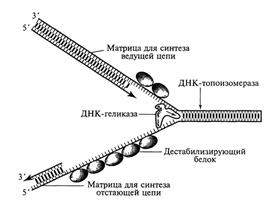
Рис. Схема образования репликационной вилки ДНК
Эти белки обладают способностью связываться только с одноцепочечными (уже раскрученными и разделенными) участками молекулы, препятствуя возникновению на них вторичных складчатых образований («шпилек») за счет случайных соединений между комплементарными нуклеотидами однонитевой структуры. Следовательно, они способствуют выпрямлению однонитевых участков молекулы, что необходимо для нормального выполнения ими матричных функций.
Быстрое расплетание ДНК с помощью хеликазы без дополнительного вращения нитей по отношению друг к другу должно приводить к образованию новых витков (узлов) на участках родительской молекулы перед движущейся вилкой репликации, создающих повышенное топологическое напряжение на этих участках. Такое напряжение устраняется еще одним белком (ДНК-топоизомеразой), который, перемещаясь вдоль двухспиральной родительской ДНК перед вилкой репликации, вызывает временные разрывы в одной из цепочек молекулы, разрушая фосфодиэфирные связи и присоединяясь к разорванному концу. Возникший разрыв обеспечивает последующее вращение нити двойной спирали, что, в свою очередь, приводит к расплетанию образующихся супервитков (узлов). Поскольку разрыв полинуклеотидной цепочки, вызванный топоизомеразой, носит обратимый характер, то разорванные концы быстро воссоединяются сразу после разрушения комплекса этого белка с разорванным концом.
На втором этапе происходит матричный синтез новых (дочерних) полинуклеотидных цепей на основе известного принципа комплементарного соответствия нуклеотидов старой (матричной) и новой цепей. Этот процесс осуществляется путем соединения (полимеризации) нуклеотидов новой цепи с помощью ферментов ДНК-полимераз нескольких типов. Следует отметить, что ни одна из известных сегодня ДНК-полимераз не способна начать синтез нового полинуклеотида путем простого соединения двух свободных нуклеотидов. Инициация этого процесса требует наличия свободного 3'-конца какой-либо полинуклеотидной цепочки ДНК (либо РНК), которая соединена с другой (комплементарной) цепочкой ДНК. Иными словами, ДНК-полимераза способна лишь добавлять новые нуклеотиды к свободному 3'-концу имеющегося полинуклеотида и, следовательно, способна наращивать эту структуру только в направлении 5'→3'.
С учетом указанного обстоятельства становится понятным асимметричный характер функционирования вилки репликации (рис. 1.9 и 1.10). Как видно из приведенных схем, на одной из матричных нитей вилки 3'→5') идет относительно быстрый и непрерывный синтез дочерней нити (ведущей, или лидирующей, цепочки) в направлении 5' →3', тогда как на другой матрице (5'→ 3') идет более медленный и прерывистый синтез отстающей цепочки короткими фрагментами (100-200 нуклеотидов), получившими название фрагментов Оказаки, и также в направлении 5'→ 3'.
Синтез ведущей и отстающей цепочек осуществляют ДНК-полимеразы разных типов. Свободный 3'-конец, необходимый для начала синтеза фрагмента Оказаки, обеспечивается короткой нитью РНК (около 10 нуклеотидов), получившей название РНК-праймера (РНК-затравки), которая синтезируется с помощью фермента РНК-праймазы. РНК-праймеры могут комплементарно спариваться сразу с несколькими участками на матричной нити ДНК, создавая условия для одновременного синтеза нескольких фрагментов Оказаки при участии ДНК-полимеразы III (рис. 1.10). Когда синтезированный фрагмент Оказаки достигает 5'-конца очередного РНК-праймера, начинает проявляться 5'-экзонуклеазная активность ДНК-полимеразы I, которая последовательно выщепляет нуклеотиды РНК в направлении 5'→ 3'. При этом удаляемый РНК-праймер замещается соответствующим фрагментом ДНК.
Последний (третий) этап рассматриваемого процесса связан с действием фермента ДНК-лигазы, который соединяет 3'-конец одного из фрагментов Оказаки с 5'-концом соседнего фрагмента с образованием фосфодиэфирной связи, восстанавливая таким образом первичную структуру отстающей цепочки, синтезируемой в функционирующем репликоне. Дальнейшая спирализация появившегося «полуконсервативного» участка ДНК (закручивание спирали) происходит с участием ДНК-гиразы и некоторых других белков.
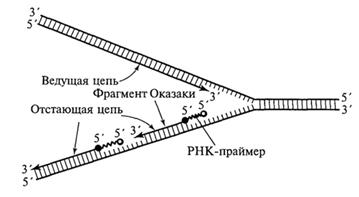
Рис. Синтез ведущей и отстающей цепей ДНК в области репликационной вилки
Полирепликонный принцип организации молекулы ДНК различных эукариот, в том числе человека, обеспечивает возможность последовательного копирования генетического материала этих организмов без одновременного раскручивания (деспирализации) всей огромной по размерам и сложно упакованной молекулы, что значительно сокращает время ее репликации. Иными словами, в тот или иной момент времени в одной группе репликонов молекулы процесс копирования может быть уже завершен объединением и спирализацией соответствующих участков, тогда как в другой группе он только начинается расплетанием двухнитевых структур.
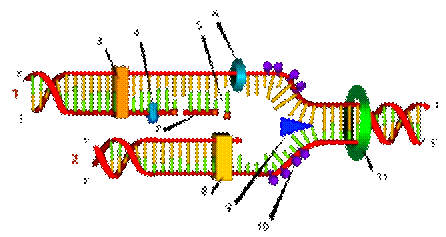
Схематическое изображение процесса репликации, цифрами обозначены: 1- запаздывающая цепь, 2 - цепь-лидер, 3- ДНК-полимераза (Polα), 4- ДНК лигаза, 5- РНК-праймер, 6- ДНК- праймаза, 7- фрагмент Оказаки, 8- ДНК-полимераза (Polδ), 9- хеликаза, 10- одиночная цепь со связанными белками, 11- топоизомераза
Ферменты, участвующие в репликации ДНК эукариот:
1.ДНК-хеликаза и дестабилизирующие белки; они расплетают двойную спираль родительской ДНК и формируют репликационную вилку.
2.ДНК-полимеразы, которые катализируют синтез полинуклеотидной цепи ДНК в направлении 3'-5, копируя в репликационной вилке матрицу с высокой степенью точности. Поскольку две цепи двойной спирали ДНК антипараллельны, в направлении 5'-3' непрерывно синтезируется лишь одна из двух цепей - ведущая; другая цепь - отстающая, синтезируется в виде коротких фрагментов Оказаки. ДНК-полимераза способна к исправлению собственных ошибок, но не может самостоятельно начать синтез новой цепи.
3.ДНК-праймаза, которая катализирует короткие молекулы РНК-затравки. Впоследствии фрагменты РНК удаляются - их заменяет ДНК.
4.Теломераза, заканчивающая построение недорепликацированых 3'-концов линейных молекул ДНК.
5. ДНК-топоизомеразы, помогающие решить проблемы кручения и спутывания спирали ДНК.
6. Инициаторные белки, связывающиеся в точке начала репликации и
способствующие образованию нового репликационного глазка с одной или двумя вилками. В каждой из вилок вслед за инициаторными белками к расплетенной ДНК сначала присоединяется белковый комплекс, состоящий из ДНК-хеликазы и ДНК-праймазы (праймосома). Затем к праймосоме добавляются другие белки и возникает «репликационная машина», которая и осуществляет синтез ДНК.
Дата добавления: 2016-03-20; просмотров: 3569;