Психофизиологический подход к определению сознания
Единого общепринятого определения сознания в психологии и психофизиологии не существует. В большинстве случаев сознание определяют через функции, которые оно выполняет. Например, нейрофизиолог X. Дельгадо (1971), автор широко известной книги «Мозг и сознание», приводит определение сознания как организованной группы процессов в нервной ткани, возникающих немедленно на предшествующие интрапсихические (вызванные внутренними причинами) или экстрапсихические (вызванные внешними причинами) события. Эта группа нервных процессов, т.е. сознание, воспринимает, классифицирует, трансформирует и координирует вызвавшие его события с целью начать действие на основе предвидения его последствий и в зависимости от наличной информации.
В других определениях подчеркиваются системность сознания, комплексность выполняемых им функций, связь с памятью (прошлым и будущим человека), привязанность к мозговому субстрату. П.В. Симонов (1987), например, особо выделяет коммуникативный аспект сознания, определяя его как оперирование знанием, способность к направленной передаче информации от одного лица к другому.
Однако для понимания физиологической природы сознания большее значение имеют существующие представления о механизмах, лежащих в основе функциональных состояний, и, в первую очередь, концепция уровней бодрствования (см. главу 3).
Сознание и уровни бодрствования.При анализе сознания как психофизиологического феномена необходимо четко разводить два его аспекта. Во-первых, сознанию соответствует определенный диапазон в существующем континууме «сон-бодрствование». Известно, что при сильном снижении уровня бодрствования развивается состояние, которое определяется как кома («без сознания»). Очевидно также, что при относительно низких уровнях бодрствования, например во сне, сознание в полном объеме своих функций не выявляется. Именно поэтому сон предлагается квалифицировать как измененное состояние сознания. Физиологическим условием проявления сознания служит состояние пассивного и активного бодрствования.
Во-вторых, в качестве самостоятельной характеристики предлагается выделять содержание сознания. Последнее непосредственно связано с психическим отражением и выполняет все функции, перечисленные в определении, данном выше.
Очевидно, что обе стороны сознания тесно связаны между собой. Так при пробуждении ото сна по мере возрастания уровня бодрствования содержание сознания становится все более насыщенным. В то же время при очень сильном эмоциональном напряжении, когда уровень бодрствования достигает наиболее высоких значений, содержание сознания начинает страдать, происходит его своеобразное «сужение». Наконец, существует словосочетание «ясное сознание», т.е. такое состояние, когда человек свободно реализует все перечисленные выше функции сознания, и принятые им решения наиболее осознаны. Есть все основания полагать, что этому соответствует особый уровень возбуждения коры больших полушарий, который именуется оптимальным.
Итак, в психофизиологии сознание понимается, в первую очередь, как особое состояние мозга, при котором только и возможна реализация высших психических функций. Другими словами, сознание – это специфическое состояние мозга, позволяющее осуществлять определенные когнитивные операции (Соколов, 1997). Выход из этого состояния приводит к выключению высших психических функций при сохранении механизмов жизнеобеспечения.
Сознание и внимание.Классик экспериментальной психологии Э. Б. Титченер определял внимание как «способность и возможность концентрировать сознание». Он выделял два уровня сознания, из который «верхний слой» относил к зоне ясного видения, а «нижний» – к зоне смутного. Сходным образом определял внимание С.Л.Рубинштейн: «поле нашего сознания не плоскостно.
Часть сознаваемого выступает на передний план в виде «фигуры» на отступающем и стушевывающемся фоне». И в когнитивной психологии внимание определяется как направленность «деятельности по переработке информации на ограниченную часть «входа», при этом существует предвнимание, представляющее «деятельность за пределами основного потока информации» (Найсер, 1976). Таким образом, избирательное внимание в психологии всегда связывалось с ограничением сознания.
Физиологически избирательное внимание проявляется в локальной активации (повышенной возбудимости) определенных участков коры в сочетании с более или менее выраженным торможением остальных областей коры больших полушарий. В зависимости от характера деятельности зона повышенной активации может перемещаться по коре больших полушарий. По некоторым данным наиболее высокому уровню сознания и избирательного внимания соответствует локализация очага повышенной активации в левой фронтальной области, а наиболее низкому – в правой затылочной. Таким образом, локальная активация коры больших полушарий при осуществлении сознательной деятельности имеет динамический характер, определяемый сигнальной значимостью действующих раздражителей и характером выполняемой деятельности (Костандов, 1983).
Нейрофизиологические основы сознания.И.П.Павлов образно представлял сознание как перемещающуюся по коре зону повышенной возбудимости – «светлое пятно сознания» на темном фоне остальной коры. В настоящее время согласно данным, полученным с использованием метода ПЭТ-томографии, зона локальной активации действительно имеет вид светлого пятна на темном фоне.
Современным аналогом представлений Павлова можно считать теорию «прожектора» (Crick, 1984). Решающая роль в ней отводится таламусу, именно он направляет поток возбуждения в кору больших полушарий. Причем процесс осуществляется таким образом, что в каждый данный момент времени только один из таламических центров находится в состоянии возбуждения, достаточном для создания в коре зоны повышенной возбудимости. Период такой высокой возбудимости длится около 100 мс, а затем усиленный неспецифическим таламическим возбуждением приток импульсов поступает к другому отделу коры. Область наиболее мощной импульсации создает центр внимания, а благодаря постоянным перемещениям потока возбуждения по другим участкам коры становится возможным их объединение в единую систему.
В качестве предполагаемого механизма, связывающего нейроны в единую систему, Крик рассматривает появление у них синхронизированных разрядов с частотой в гамма-диапазоне 35 – 70 Гц. Синхронизация нейронной активности является механизмом объединения клеток в ансамбль. Таким образом, согласно теории Крика, нейронные процессы, оказывающиеся в центре гипотетического «луча прожектора», определяют содержание нашего сознания в текущий момент времени, а функцию управления лучом прожектора выполняет таламус, создавая в разных зонах коры длящееся около 100 мс неспецифическое возбуждение (локальную активацию).
По описанным выше представлениям возникновение сознания становится возможным только при достижении определенного уровня возбуждения коры больших полушарий. Идея порога возбуждения как условия возникновения сознания, не может, однако, объяснить многие аспекты функционирования сознания. По этой причине еще в 80-е годы был предложен другой нейрофизиологический механизм возникновения сознания, получивший название теории повторного входа (Эделмен, 1981). В этой теории в качестве центрального механизма возникновения сознания рассматривается механизм повторного входа возбуждения. Имеется в виду, что возбуждение, возникшее в группе нейронов первичной зоны коры, возвращается в ту же нейронную группу после дополнительной обработки информации в других нервных центрах или поступления новой информации из внешней среды, а также из долговременной памяти. Этот повторный вход дает возможность сопоставлять имевшиеся раньше сведения с изменениями, которые произошли в течение одного цикла обработки информации. Объединение двух потоков информации (первичного и вторичного) составляет один цикл или повторяющуюся фазу активности сознания. Прохождение одного цикла возбуждения осуществляется за 100 – 150 мс. Повторяющиеся циклы создают нейрофизиологическую «канву» сознания.
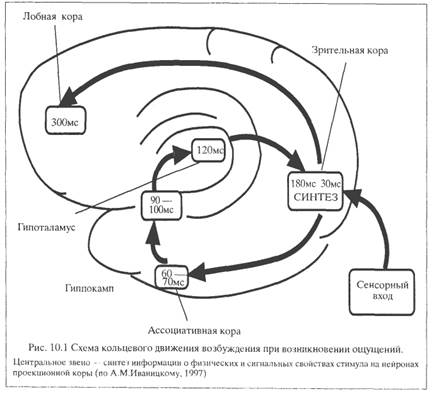
Представления Дж. Эделмена получили дальнейшее развитие, в частности, в концепции А.М.Иваницкого (1997). Центральное место в этой концепции занимает понятие информационного синтеза (см. главу 5.3), предусматривающее объединение информации о физических и семантических особенностях стимула. Информация о физических свойствах стимула поступает по сенсорно-специфическим путям, а информация о значимости стимула извлекается из памяти. Синтез этих двух видов информации (текущей и извлекаемой из памяти) обеспечивает возникновение ощущения. Этот процесс осуществляется с периодом квантования приблизительно в 100-180 мс. Опознание стимула происходит с участием лобных областей мозга, приблизительно через 300 мс от момента предъявления. Последовательное поступление информации от рецепторов приводит к повторному движению возбуждения по указанному кольцу, обеспечивая постоянное сопоставление сигналов, приходящих из внешней и внутренней среды.
В последнее время при обсуждении нейрофизиологических основ сознания большое внимание уделяется высокочастотной биоэлектрической активности мозга (Данилова, 1998). Ведущая роль в возникновении сознания приписывается высокочастотным составляющим спектра ЭЭГ в диапазоне от 35Гц до 120Гц. Сознание определяют как продукт синхронизации нейронной активности в указанном частотном диапазоне. Предполагается, что когерентная электрическая активность достаточно большого числа нейронов создает необходимое и достаточное условие для явления сознания даже в отсутствии сенсорной стимуляции, как, например, во сне.
В этом контексте большое значение приобретает временная последовательность, объединяющая активность множества нейронов в единый пространственно-временной паттерн. Принцип «временного связывания» элементарных символов (нейронов ) в более сложные структуры на основе корреляции их сигналов во времени рассматривается как основополагающий. Причем особое значение придается частоте 40Гц, которая оптимальным образом обеспечивает синхронизацию импульсной активности нейронов, участвующих в образовании образов и поддержании сознания.
В заключении следует отметить, что приведенные выше представления о нейрофизиологических механизмах сознания отнюдь не исчерпывают всего многообразия в изучении этой проблемы (см. Данилова, 1998). Тем не менее они весьма иллюстративны в своем многообразии, выдвигая на первый план разные нейрофизиологические механизмы: перемещение по коре фокуса повышенной возбудимости, возврат возбуждения и синхронизацию высокочастотной активности нейронов.
Дата добавления: 2016-03-10; просмотров: 837;