Критерии соответствия
К 1980 году исследователи памяти уже достаточно хорошо разбирались, чему можно верить, а чему нет. Стало, например, ясно, что не существует уникальных молекул памяти. Сложилось общее мнение, что наиболее перспективна модель Доналда Хебба, в которой предполагалось изменение эффективности связей между нервными клетками, вероятно, в результате образования новых или роста существующих синапсов, и этим объяснялась перестройка функциональных отношений между нейронами. Модификации хеббовского типа, по-видимому, можно было бы выявлять нейрофизиологическими или биохимическими методами. Однако ни у кого не было уверенности, что какой-либо специфический биохимический процесс, кроме самого общего процесса белкового синтеза, можно однозначно связать с формированием следов памяти; ученые стали проявлять большую осторожность при оценке новых сообщений на эту тему. Что же такое в конце концов биохимия памяти и как проверить правильность предлагаемых ответов на этот вопрос? Иными словами, какой ответ был бы убедительным и для биохимиков, и для психологов? Каким требованиям должен отвечать эксперимент, чтобы изучаемый процесс можно было считать необходимым, достаточным и, быть может, даже специфичным для образования следов памяти?
На этот вопрос я попытался ответить в начале восьмидесятых годов [1]. К тому времени уже не составляло труда обнаружить биохимические изменения в мозгу обучавшегося животного; трудность заключалась в доказательстве того, что они действительно связаны с процессом запоминания. Я чувствовал, что не могу разумно планировать эксперименты, не имея критериев для оценки их результатов, а без этого я не знал бы, как двигаться дальше. Я решил, что любое изменение, которое можно будет рассматривать как часть следа памяти (энграммы), должно соответствовать критериям, описанным ниже1.
*1) Я несколько изменил формулировку этих критериев и порядок их обсуждения по сравнению с тем, как они были представлены в оригинальной статье, но это не затрагивает их сути. Я не хочу переписывать историю.
Первый критерий
Предполагаемый фактор (система, вещество, скорость его образования или обновления) в локализованном участке мозга должен претерпевать количественные изменения в процессе формирования следов памяти.
Если образуются новые или видоизменяются существующие синапсы, то в их химических или структурных компонентах должны появляться изменения, доступные для выявления биохимическими или микроскопическими методами (например, повышенное содержание синаптических мембранных белков, изменение размеров определенных синапсов или увеличение их числа). Однако если бы я выявил такое изменение в период обучения животного, но не убедился в его соответствии всем последующим критериям, я бы недалеко ушел от экспериментов шестидесятых годов, которые критиковал в предыдущей главе. То или иное изменение может быть действительно необходимым, но это еще не значит, что оно достаточно или специфично. Тем не менее первый критерий, безусловно, имеет фундаментальное значение для любой материалистической модели памяти. Он предполагает также условие локализации, т. е. изменения не могут быть распределены по всему мозгу - они должны быть сосредоточены в какой-то определенной области. Это, разумеется, возвращает нас к старому спору о возможности локализации энграмм (к этой теме я вернусь позднее; значительная часть двух последних глав будет посвящена вопросу о локализации памяти во времени и пространстве).
Обратите внимание, что первый критерий не включает таких условий, как направление и величина изменений. Вполне возможно представить себе «отрицательное» кодирование воспоминаний путем снижения уровня какого-то вещества или процесса, хотя на практике почти цсе исследователи пытаются выявить его повышение. А что же с величиной изменения?
Если что-то запомнилось вдвое лучше или вместо одного события или предмета запомнились два, значит ли это, что и изменение выражено вдвое сильнее? Совсем не обязательно, ибо в наших опытах мы измеряем память в условных единицах, которые сами же изобрели. При этом мы не можем знать, соответствуют ли наши масштабы тем, которыми пользуются сами животные, формируя следы памяти. Например, я могу обучить цыпленка не клевать горькую хромированную бусину, потом горькую красную и, наконец, горькую синюю бусину. Однако цыпленок не обязательно помнит их как три отдельных предмета. Скорее всего он вместо этого прибегает к иной, более рациональной стратегии, производя обобщение: «Все предметы определенного размера, независимо от цвета, имеют горький вкус, и их следует избегать». В этом случае он будет помнить один предмет, а не три.
Второй критерий
Изменение должно совпадать во времени с формированием следов памяти.
Очевидно, что следы образуются не мгновенно, словно нажатием кнопки, а на протяжении нескольких часов после запоминаемого события. В этот период изменяется форма сохранения следа. По крайней мере в случае декларативной памяти (глава 4) имеет место переход, занимающий несколько минут или часов, от первоначальной лабильной и кратковременной фазы к долговременной стабильной памяти.
Можно представить себе разные способы такого перехода (рис. 9.1). Одним из крайних вариантов может быть непрерывный процесс, в котором некая цепь биохимических реакций в определенной группе клеток неизбежно приводит от ранней, неустойчивой фазы к фиксированному конечному состоянию, как при затвердевании клея или проявлении фотоснимка. Другой вариант допускает возможность двух (или нескольких) более или менее независимых параллельных процессов, сопровождающихся изменениями электрических свойств и чувствительности какой-то группы нейронов, которые могут в первые минуты «кодировать» память, а затем постепенно отключаться. Между тем, если воспоминание достаточно важно, чтобы стоило сохранять его долгое время, могут потребоваться такие биохимические процессы, как реконструкция синапсов, которые будут постоянно представлять память в другой группе нейронов, возможно даже в другой части мозга. Эти два типа процессов - последовательная и параллельная обработка воспоминаний - служат, разумеется, крайними примерами; есть множество промежуточных вариантов [2], и становится все труднее планировать эксперименты, в которых можно было бы четко провести различие между ними.
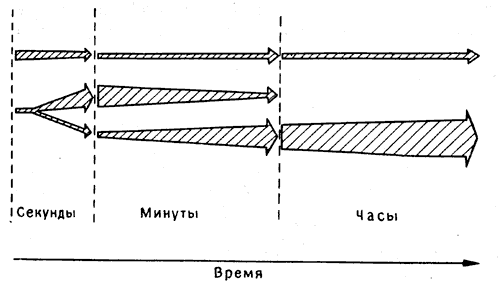
Рис. 9.1. Кратковременная и долговременная память. Две модели перехода от кратковременной памяти к долговременной. Вверху показан вариант с последовательным, линейным переходом от фазы к фазе. Внизу - вариант с двумя независимыми процессами. Первоначальная активация клеток служит сигналом как для запуска механизмов кратковременного запоминания, так и для образования и последующего укрепления долговременных энграмм. При блокировании второго из этих процессов воспоминание утрачивается по мере угасания кратковременных следов. Имеющиеся данные говорят скорее в пользу нижнего варианта.
Тем не менее весь опыт изучения человеческой памяти указывает именно на такое подразделение памяти на кратковременную и долговременную. Результаты исследований, проведенных на таких больных, как X. М., показывают, что гиппокамп, не участвуя в проявлениях долговременной памяти, все же играет важную роль в переходе от кратковременной к долговременной фиксации ее следов. Но как бы ни работал мозг - путем последовательной или параллельной обработки информации (в дальнейших главах этому будет уделено больше внимания) - в обоих случаях с памятью связаны определенные клеточные процессы, и мы не можем не пытаться дифференцировать их экспериментальным путем.
Третий критерий
Стресс, двигательная активность и другие процессы, сопровождающие научение, не должны приводить к структурным или биохимическим изменениям, если при этом не образуются следы памяти.
Теоретически этот критерий очевиден, но его очень трудно проверить на практике. Если обучение животного в эксперименте невозможно без стресса, двигательной активности и т. п., то возможны ли стресс, двигательная активность и другие процессы без научения? Имеет ли смысл попытка такого разделения, и можно ли представить себе такой вариант в высшей степени редукционистского эксперимента, который позволил бы осуществить такое разделение? Не гонимся ли мы за чем-то неосуществимым ни экспериментально, ни теоретически? Вправе ли я вообще ставить этот вопрос, учитывая, сколько энергии я потратил на критику редукционизма с теоретических позиций?
Но именно потому, что на протяжении последних двух десятилетий я потратил столько времени в попытках получить результаты, удовлетворяющие обсуждаемому критерию, нелишне повторить, что следование редукционистской методологии в стратегии исследований (т. е. стремление стабилизировать изучаемый мир, манипулируя лишь одной переменной и сохраняя остальные по возможности стабильными) - это обычно единственный способ постановки эксперимента, позволяющий сделать четкие выводы. Ошибки случаются, когда переоценивают значимость таких выводов, забывая об искусственно ограниченных условиях эксперимента и вместе с тем допуская, что в реальной жизни, за стенами лаборатории, могут действительно иметь место изменения одной переменной; что очень просто экстраполировать искусственные условия лабораторной изоляции на сложно переплетенные условия реального мира. Именно этот прием, посредством которого редукционистская методология превращается в редукционистскую философию, столь популярен среди молекулярных биологов и части генетиков, но, к счастью, значительно реже используется психологами или нейробиологами. (Именно психологи подвергли наибольшему сомнению теоретическую обоснованность моего третьего критерия.)
Если не считать искусственных ситуаций, переменные находятся в непрерывном взаимодействии, и это взаимодействие носит не просто аддитивный характер. Чаще всего приводят пример из области генетики: там долго бытовало весьма наивное представление, что физиологию и поведение организма (его фенотип) можно условно разделить на два компонента, один из которых зависит от генов, а другой от окружающей среды. Таким образом, считалось, что фенотип организма практически полностью определяется совокупностью этих двух, очевидно независимых, компонентов. На самом деле, разумеется, взаимодействие между генами и факторами окружающей среды в процессе развития носит далеко не линейный характер, и попытки разделить фенотип на генетический и средовой компоненты обречены на неудачу. Пэт Бейтсон находит здесь аналогию с выпечкой пирожного. Смешивают разнообразные компоненты - муку, молоко, масло, сахар, специи, яйца и т. д., а потом смесь подвергают тепловой обработке. Каждый из этих компонентов необходим, чтобы готовое изделие имело желаемый вкус, однако бессмысленно задаваться вопросом, в какой мере этот вкус будет зависеть от муки, от яиц, от времени и температуры выпечки: в процессе смешивания и нагревания произошли качественные изменения составляющих компонентов.
Упрощенные концепции аддитивного взаимодействия генов и факторов среды хотя и фигурируют еще в стандартных учебниках генетики (часто даже без упоминания о его значении для здоровья человека), на самом деле очень мало связаны с тем, что происходит в реальной жизни. То же относится к исследованиям памяти. Если нельзя показать, что животное обучилось, иначе как по изменению поведения, и это изменение можно вызвать только определенной формой стресса или принуждения, то сопутствующие обучению биохимические изменения должны включать также изменения в ответ на стресс, в том числе эффекты нейромодуляторов, обсуждавшиеся в предыдущей главе. Тем не менее в том искусственном мире, который создается в лаборатории, можно и должно выделять переменные величины, каким-то образом связанные с фактами реальной жизни. У нас нет иного способа узнать что-либо о выпечке пирожного, кроме наблюдения за тем, что происходит, если исключать из смеси отдельные компоненты, изменять температуру или время приготовления и т. д. ...
Четвертый критерий
Если клеточные или биохимические изменения ингибируются в период, когда должно было происходить формирование энграмм, то последнее блокируется и у животного возникает амнезия, и наоборот.
Очевидно, что это логическая необходимость, и она составляет основу вмешательства в процессы памяти при использовании ингибиторов белкового синтеза, о которых шла речь в предыдущей главе. Однако это все не так просто, поскольку на практике ни один из таких ингибиторов нельзя считать «волшебной пулей», попадающей в единственную мишень и не дающей так называемых «побочных эффектов»; поэтому результаты экспериментов с ингибиторами обычно не дают однозначных результатов. Последние слова, которые я добавил к формулировке данного критерия, это что-то вроде барочного росчерка. Допустим, животное тренируют на выполнение задачи, что в обычных условиях приводит к ее усвоению, но в данном эксперименте этого не происходит, так как научение каким-то способом блокируется. В этом случае изучаемый биохимический процесс, если он действительно специфически связан с образованием следов памяти, должен отсутствовать, когда отсутствует запоминание. Эксперименты, основанные на этом критерии, описаны в следующей главе.
Пятый критерий
Удаление участка, в котором происходят биохимические, клеточные или физиологические изменения, должно препятствовать образованию следов памяти (или вспоминанию) в зависимости от того, когда по отношению к времени тренировки произведено удаление.
Этот логический аналог двух предыдущих критериев тоже может казаться очевидным. Если изменения связей, формирующие след памяти, локализованы в отдельной небольшой группе клеток, а не распределены в мозгу диффузно, то удаление этих клеток приведет к исчезновению энграммы или помешает ее формированию. Поскольку в эксперименте довольно просто создать в мозгу небольшие локализованные «дырки», не вызывая общего поражения, нетрудно проверить справедливость утверждения, что какой-то участок является тем самым носителем энграммы, который определяет данное поведение. Если соответствующее повреждение приводит к амнезии или неспособности к обучению, это может подтвердить локализацию энграммы. Однако этот вывод отражает статичный и механистический взгляд на способ фиксации воспоминаний в мозгу. Если же процесс хранения следов более динамичен и в нем участвует много различных участков, такой эксперимент ничего не даст. Кроме того, игнорируется пластичность мозга (при удалении одного участка его функцию может принимать на себя другой участок); этот вопрос исследуется в экспериментах, описанных в главе 11. Наконец, никогда не следует забывать о свойственной всем травмирующим экспериментам двойственности: вспомним пример Ричарда Грегори с радиоприемником и подавляющим шумы транзистором.
Шестой критерий
При нейрофизиологической регистрации сигналов из мест изменения на клеточном уровне должны выявляться изменения в электрических ответах нейронов во время и (или) после формирования следов памяти.
Если верна гипотеза Хебба о хранении следов памяти в форме измененных синаптических связей, то обучение должно сопровождаться изменением электрической активности нейронов, соединенных изменившимися синапсами. Данная мною формулировка этого критерия предполагает, что начинать надо с поиска изменений на биохимическом и клеточном уровнях и уже на этой основе выявлять нейрофизиологические изменения, т. е. нейрофизиология в известном смысле рассматривается просто как побочный продукт биохимических и клеточных изменений. Разумеется, эта точка зрения отражает лишь мои личные склонности как не до конца перестроившегося нейро-химика. На практике нейрофизиология может с таким же успехом вести (а в таких важных случаях, как эксперименты на аплизии или изучение долговременной потенциации, и в самом деле вела) за собой биохимию и клеточную биологию, помогая находить клетки, чьи электрические, а значит, и биохимические свойства изменяются при научении. Я не имею в виду, что биохимия первична или в редукционистском смысле более фундаментальна, чем физиология; я утверждаю только, что измененная биохимия претворяется в измененную физиологию точно так же, как и в измененное поведение.
Эти шесть критериев формировали мои собственные исследования, начиная с восьмидесятых годов. Я пытался выявить биохимические, морфологические и физиологические изменения в отдельных областях мозга цыплят в период от нескольких минут до нескольких часов после тренировки в выполнении простой задачи с целью 1) показать, что эти изменения связаны именно с образованием следов памяти, а не с какими-либо побочными аспектами обучения; 2) показать, что блокада этих изменений предотвращает запоминание, и наоборот; а также 3) изучить последствия удаления изменяющихся участков мозга до и после обучения. Результаты использования этих критериев я опишу в двух последующих главах, единственными героями которых будут цыплята; а сейчас речь пойдет не о моих собственных экспериментах, а о важнейших работах других исследователей, проводившихся в последнее десятилетие.
Новые модели
Все точные критерии и четкие теории в мире ничего не значат без хороших экспериментальных модельных систем. В восьмидесятые годы по мере исчезновения прежних сомнений в правильном выборе изучаемых объектов и проводимых тестов ученые начали приходить к единому мнению, что лучше использовать небольшое число моделей, причем каждая группа исследователей отстаивала свою новую версию «идеального» организма. Правда, виды очень многообразны, и они обучаются самым разным вещам, совсем не похожим на те экспериментальные тесты, которыми пользовались первые поколения зоопсихологов. Эти психологи, однако, создали некоторый задел для своих последователей, видевших свое прямое призвание в разработке биохимии, нейрофизиологии и клеточной биологии научения и памяти. Те, кто работал с позвоночными, проявляли тенденцию уйти от прежних лабиринтов и скиннеровских клеток, чтобы заняться выработкой очень простых классических условных рефлексов (например, изменения частоты сердечных сокращений или моргания) у кроликов, у которых можно достаточно точно картировать нервные цепи.
Я уже попутно упоминал ряд других, более экзотических моделей. Например, в Иерусалиме Ядин Дудаи использовал поведенческие и биохимические мутации у плодовой мушки. Для него и некоторых других нейробиологов дрозофила стала таким же обычным объектом изучения, каким была в нашем столетии для генетиков. Доводы этих ученых основывались на том факте, что, как правило, любая специфическая точечная мутация приводит к изменению или отсутствию одного белка в развивающемся организме. Таким белком может быть, например, фермент или компонент мембраны. У мутанта, дефектного по признаку обучаемости или памяти, причиной дефекта может быть отсутствие специфического белка или нарушение его функции. Если установить, какой белок отсутствует, то можно получить ключ к расшифровке его роли в образовании энграмм. В этом смысле изучение мутаций несколько напоминает применение ингибиторов для блокирования определенных обменных процессов и обладает всеми достоинствами и недостатками последнего метода, которые я обсуждал в предыдущей главе и в разделе «Четвертый критерий». Исследования на дрозофилах не решили проблему памяти, но они, несомненно, содействовали пониманию ее биохимических механизмов. Одним из важнейших результатов этих работ явилась демонстрация сходства молекулярных процессов, лежащих в основе формирования следов памяти у плодовой мушки и у других, более крупных и обычных лабораторных животных. Эти результаты подтверждают существование подлинно универсальных биохимических основ нервной пластичности.
Другим очень популярным объектом для изучения памяти стали моллюски; о причинах этого говорилось в главе 7. Особенно важно наличие у них крупных нейронов и легко доступной для исследования нервной системы. Моделями могут служить несколько интересных видов наземных улиток, но наибольшей популярностью (если не у гастрономов, то у нейробиологов) пользуются гигантские морские слизни, такие как Hermissenda.
Однако два самых успешных и наиболее часто цитируемых модельных исследования 80-х годов были проведены на брюхоногом моллюске аплизии (клеточные механизмы кратковременного научения и аналогичных процессов) и на гип-покампе - особом участке в мозгу млекопитающих (феномен так называемой долговременной потенциации). Результатам этих экспериментов и посвящена оставшаяся часть главы.
Привыкание у аплизии - «обучение в блюдце»
Спросите любого специализирующегося в нейронауках выпускника университета, какой организм больше всего использовался в клеточной биологии научения, и вы скорее всего услышите в ответ: аплизия. А на вопрос об исследователе, сыгравшем ключевую роль в этой программе работ, вам почти наверняка назовут имя Эрика Кэндела, профессора Фонда Говарда Хьюза в Нью-Йоркском колледже терапевтов и хирургов, автора одного из самых известных в последнем десятилетии пособий по нейронаукам [3] и плодовитого соавтора других книг, блестящего и неутомимого пропагандиста аплизии как идеального объекта для изучения памяти и редукционизма как методологической и философской концепции ее познания (Кэндел был столь ярым приверженцем редукционизма как философии и одновременно методологии, что однажды выступил перед аудиторией психблогов с лекцией на тему «Психотерапия и отдельный синапс» [4]).
Кэндел, получивший образование как психиатр, в шестидесятые годы какое-то время работал с аплизиями в Париже вместе с Ладиславом Тауком и имел возможность оценить перспективность этого моллюска при изучении вначале таких кратковременных процессов, как привыкание. В последующую четверть века этот организм стал главным объектом его самостоятельных исследований в Нью-Йорке, где в возглавляемой им лаборатории Колумбийского университета выросло несколько поколений специалистов-нейробиологов. Несомненно, Кэндел и его школа внесли важный вклад в нейробиологию кратковременных проявлений памяти как в экспериментальном плане, так и в смысле повышения престижа того раздела физиологии, к которому после его упадка в шестияесятые годы многие относились весьма сдержанно. Далеко не просто складывались личные отношения между учеными, работавшими с аплизией, и между всей этой группой и другими группами,, которые использовали иные модели, в частности с Дэном Элконом и его коллегами по работе с Hermissenda в Вудс-Хоуле (см. главу 7). Одно время трения стали столь заметными, что побудили автора научно-популярных работ Сюзан Олпорт посвятить им отдельную книгу [5]. Тем не менее общее направление исследований Кэндела и теоретические предпосылки, разработанные им в 70-х и 80-х годах, до недавнего времени не встречали особых возражений. Однако результаты, полученные в последние несколько лет в его собственной и в других лабораториях, побуждают расширить прежний несколько упрощенный редукционистский подход к интерпретации получаемых данных. Чтобы дать представление о существе этой критики, мне придется сначала рассмотреть редукционистскую концепцию Кэндела в ее наиболее выраженной форме.
В главе 7 уже говорилось, почему исследования на аплизии приобрели стратегическое значение для нейробиологии некоторых важных форм памяти. У этого животного очень несложный поведенческий репертуар, включающий различные виды научения, а их центральную нервную систему сравнительно нетрудно картировать, так как она состоит не более чем из 20 000 нейронов, сгруппированных в ряд обособленных ганглиев, и среди этих нейронов есть очень крупные клетки, легко распознаваемые индивидуально у всех особей. Ключевым моментом в подходе Кэндела было изучение простейших поведенческих реакций у интактного животного. Речь идет об элементарном рефлексе - втягивании жабры и сифона - с явлениями привыкания и сенситизации. Эту форму поведения или ее нервный механизм можно «отделить» от других поведенческих реакций путем последовательного уменьшения группы изучаемых в эксперименте нейронов. Предельным случаем будет наблюдение над двумя определенными нейронами, которые можно отпрепарировать, а затем вызвать образование синаптической связи в лабораторной чашке. По мнению Кэндела, взаимодействие таких нейронов и их реакции на добавляемые извне нейромедиаторы можно считать моделью памяти для рефлекса в ее крайне редуцированной форме. Каковы же доводы в пользу этого?
Рассмотрим сначала сам рефлекс. Аплизия дышит с помощью жабры, которая размещается в полости на верхней (дорсальной) стороне тела. В задней части этой полости имеется вырост в виде трубки - сифон. Если прикоснуться к области вокруг жабры или сифона, они втягиваются в полость: это простой защитный рефлекс. Нервный механизм такого рефлекса включает небольшое число (около 50) сенсорных нейронов, реагирующих на тактильное раздражение упомянутой области. Эти сенсорные нейроны контактируют примерно с 20 моторными нейронами как непосредственно, так и через промежуточные нейроны (интернейроны). Моторные нейроны расположены в абдоминальном (брюшном) ганглии и в свою очередь образуют синапсы на мышечных волокнах, втягивающих сифон и жабру. Эта относительно простая цепь схематически показана на рис. 9.2. Однократное раздражение поверхности тела около жабры или сифона приводит к их втягиванию, а при многократной стимуляции наступает привыкание; реакция на повторные стимулы постепенно ослабевает вплоть до ее полного исчезновения на некоторое время. Привыкание можно снять (дегабитуация, или сенситизация) сильным раздражением другого участка тела, например «хвоста», что приводит к полному восстановлению первоначальной реакции. Такие изменения рефлекса носят кратковременный и довольно неспецифический характер, и их следует рассматривать как формы неассоциативного научения. Наряду с этим, как показал Кэндел, возможна и выработка классического условного рефлекса, когда безусловным стимулом служит резкое воздействие на хвостовую область, а условным - слабое тактильное раздражение сифона. Слабая стимуляция обычно приводит и к слабо выраженному втягиванию сифона, тогда как после закрепления условного рефлекса такое же слабое раздражение вызывает сильную реакцию. Этот эффект сохраняется довольно долго, и поскольку здесь существует специфическая связь между стимулом и реакцией, его можно считать формой истинного ассоциативного научения.
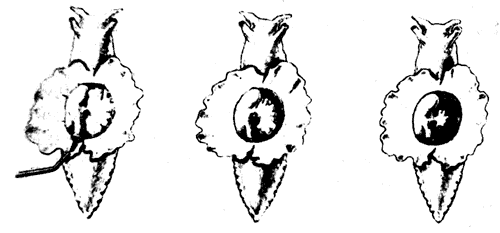
Рис. 9.2а. Рефлекс втягивания жабры и сифона у аплизии. Показано интактное животное. На сифон через трубку направлена струя воды (слева), это приводит к втягиванию сифона и фабры (в середине и справа).
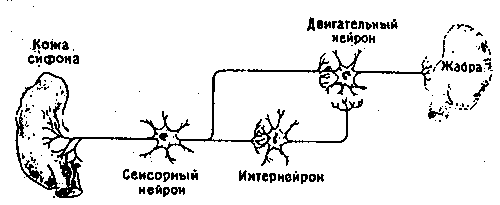
Рис. 9.2b. Схема нервных связей в «редуцированном» препарате.
Кэндел и его сотрудники занялись поисками нервных путей, участвующих в реакции втягивания жабры и сифона. Они использовали простые классические методы нейрофизиологии, а дальнейшая стратегия состояла в последовательном уменьшении числа нейронов. Для более точного контроля и количественной оценки реакции моллюсков обездвиживали, прикрепляя к пластинке, а для стандартизации тактильных стимулов применяли струю воды, направляемую через остроконечную трубку. Сокращения жабры можно было также прямо регистрировать с помощью фотоэлемента. Выявив нервную сеть, которая обеспечивала рефлекторную реакцию, исследователи смогли перейти ко второму вопросу: какая часть этой сети ответственна за привыкание? Не происходят ли при этом изменения в каких-то определенных клетках или синапсах? Этот вопрос, очевидно, имеет отношение к описанному выше первому критерию. Но, поскольку группа Кэндела состояла из нейрофизиологов, а не биохимиков, исследователи начали с изучения электрических свойств клеток, что соответствует моему шестому критерию.
Однако для получения ответа требовалось еще одно упрощение изучаемой системы, в результате которого живые аплизии превращались в неактивные, удобные для манипуляций «препараты». Животное можно вскрыть, обнажив абдоминальный ганглий, или даже полностью изолировать этот ганглий вместе с нервами, участком кожи и жаброй. Проведя такую операцию, Кэндел мог считать, что исключил все посторонние источники сигналов - другие периферические нервы, нейро-модуляторы, циркулирующие в крови и т. п. При этом он получил возможность выявлять крупные клеточные тела моторных нейронов и, как говорилось в главе 7, повторно идентифицировать «те же самые» клетки при смене одного животного другим (рис. 9.2). В результате операции живой моллюск становился чем-то вроде компьютерной схемы, и ученые могли исследовать ее свойства как электротехники, получившие новое оборудование и пытающиеся понять принципы его работы. В такой системе осязательный стимул для поведенческих реакций можно заменить его нейрофизиологическим аналогом, т. е. прямым электрическим раздражением нервов, идущих к сенсорным нейронам. Точно так же и мышечную реакцию - втягивание жабры и сифона - можно вызывать прямой стимуляцией нервов, отходящих от моторных нейронов к жаберной мускулатуре.
На следующем этапе работы изолированные и «упрощенные» препараты использовались для того, чтобы выяснить локализацию механизмов привыкания, т. е. найти те звенья нервной цепи, которые начинают слабее отвечать на повторное раздражение. К середине семидесятых годов стало ясно, что ни подходящие к сенсорным нейронам, ни отходящие от двигательных нейронов нервы не обладают свойствами, позволяющими объяснить привыкание, так как их электрические ответы не уменьшаются. Следовательно, клетки, ответственные за привыкание, должны занимать промежуточное положение в сенсорно-моторной цепи абдоминального ганглия. И действительно, регистрация электрических сигналов от моторных нейронов в этом ганглии в период привыкания выявила постепенное уменьшение их частоты. Из этого был сделан вывод, что «место» привыкания лежит между сенсорными и моторными нейронами [6].
Однако даже в таком крайне упрощенном препарате имеется множество нервных цепей и многие тысячи клеток. Сенсорные нейроны, например, имеют прямые и опосредованные связи с моторными нейронами. Прямые связи обеспечиваются синапсами между аксоном сенсорного нейрона и дендритом или телом моторного нейрона (моносинаптический путь); в непрямом, полисинаптическом пути сначала сенсорный нейрон устанавливает синаптическую связь с интернейроном, а тот в свою очередь образует синапс с моторным нейроном. (Здесь следует ввести еще два соотносящиеся друг с другом термина. Когда одна клетка воздействует на другую непосредственно путем модификации синапса, образуемого ею на второй клетке, говорят о гомосинаптическом эффекте; если же действие одной клетки на другую модулируется третьей клеткой, имеющей синапсы с двумя первыми, такой эффект называют гетеросинаптическим.)
Анализ активности моторных нейронов аплизии после сенсорной стимуляции показывает, что они реагируют как непосредственно, т. е. моносинаптически, так и полисинаптически, через посредство интернейронов. На просьбу указать наиболее вероятное место синаптической пластичности теоретики скорее всего выберут интернейроны, поскольку они, очевидно, могут получать и модулировать сигналы, поступающие по множеству различных входов, прежде чем направить их по различным выходным путям. Простая модель научения, предложенная Хеббом и представленная на рис. 6.1, требует участия трех нейронов. Иными словами, если она верна, то предполагает гетеросинаптическое взаимодействие. Однако, к удивлению многих, в начале восьмидесятых годов группа Кэндела показала, что элемент, ответственный за привыкание, чрезвычайно прост: это прямая синаптическая связь между сенсорным и моторным нейронами; в частном случае - синапс между сенсорным нейроном и одним определенным очень крупным моторным нейроном, причем связь здесь моносинаптическая с модуляцией гомосинаптического типа.
Постепенное упрощение препарата до изолированной нервной цепи подготовило почву для его предельной редукции: сотрудник Кэндела Самюэл Шахер выделил два специфических нейрона - сенсорный и двигательный - и поместил их вместе в чашку Петри, т. е. перенес в условия тканевой культуры. Уже давно известно, что в этих условиях нейроны, подобно другим клеткам, много дней и даже недель остаются живыми, если поддерживать нужную температуру, аэрировать среду и снабжать клетки глюкозой и другими необходимыми веществами. При этом многие виды культивируемых клеток продолжают делиться, а нейроны хотя и не делятся, но растут и образуют аксоны, дендриты и даже синаптические соединения. В культурах Шахера сенсорные нейроны формировали синапсы на моторных нейронах, и их электростимуляция вызывала электрическую реакцию последних. При многократном раздражении сенсорной клетки ответ моторного нейрона постепенно затухал, т. е. развивалось привыкание [7]. В конце концов в лаборатории Кэндела было получено то, что он описал как изолированный синапс, «обучающийся в блюдце». Фактически это был полный триумф редукционистской стратегии, которой следовала группа Кэндела. Это, казалось, оправдывало его претензию на постановку такой цели, как расшифровка «клеточного алфавита» научения. Сенсорно-моторный синапс, безусловно, может считаться одной из букв такого алфавита.
Все, о чем шла речь до сих пор, относится в основном к области нейрофизиологии. А что можно сказать о биохимических механизмах реакций на изученных Кэнделом уровнях клеточной организации? Если привыкание происходит вследствие ослабления постсинаптической реакции в отдельном синапсе, то по логике вещей оно должно быть результатом либо пресинаптического, либо постсинаптического процесса или, разумеется, их сочетания. Например, может постепенно уменьшаться количество нейромедиатора, выделяемое пресинаптической клеткой, или происходить изменение рецепторов на постсинаптической стороне, так что они будут слабее реагировать на прежние дозы медиатора; возможно и одновременное участие обоих механизмов. Вопрос об относительной роли пре- и постсинаптической пластичности в последние годы стал одним из важных предметов спора, но большинство теоретиков склонно было приписывать пластичность в основном постсинаптической стороне.
Первой задачей нейрохимического исследования для группы, работавшей с аплизией, стала идентификация медиатора, передающего сигналы в синапсе. Им оказалось широко распространенное вещество серотонин, или 5-гидрокситриптамин. К середине семидесятых годов сотрудники группы установили, что в период привыкания в изолированном ганглии происходит неуклонное снижение секреции серотонина пресинаптическими окончаниями сенсорных клеток, тогда как чувствительность постсинаптических рецепторов этого вещества не изменяется. Пониженная секреция серотонина сопровождалась также изменением свойств пресинаптической мембраны, в особенности уменьшением трансмембранного переноса кальция внутрь клетки (биохимическое значение этого станет понятным позже). В параллельных экспериментах было показано, что сенситизация (т. е. эффект, в известном смысле противоположный привыканию; см. главы 6 и 7) тоже связана с пресинаптическими процессами, но здесь уже необходимо усиление секреции серотонина и притока кальция в клетки. Тот факт, что оба процесса - и привыкание, и сенситизация - происходят с участием пресинаптических механизмов, оказался довольно неожиданным для тех, кто моделирует нервные функции.
Десятилетие спустя были проведены аналогичные эксперименты на клетках в культуре, и они дали те же результаты. На этот раз для «обучения» не требовалось даже двух клеток, реакция полностью воспроизводилась при воздействии серотонином на изолированный моторный нейрон. Дальнейшее упрощение трудно придумать!
В следующей главе я гораздо подробнее остановлюсь на биохимии этих процессов в контексте моих собственных экспериментов. Сейчас я не буду вдаваться в детали, а хочу лишь подчеркнуть то, что Кэндел объясняет рефлекс, привыкание и сенситизацию, переходя к все более простым системам. Сначала он сводит сложную форму поведения интактного организма - втягивание сифона и жабры - к реакции нервной цепи, в которой в результате взаимодействия всего лишь двух клеток развивается привыкание, а потом интерпретирует электрическую (т. е. физиологическую) реакцию синапсов как каскад биохимических процессов в пресинаптическом нейроне.
Дата добавления: 2016-03-05; просмотров: 1239;