ГЛАВА 67. АДЕНИЛАТЦИКЛАЗНАЯ СИСТЕМА
Генри Р. Боурн (Henry R. Bourne)
Циклический 3`5`-монофосфат (циклический АМФ) действует в качестве внутриклеточного вторичного медиатора для множества разнообразных пептидных гормонов и биогенных аминов, лекарственных средств и токсинов. Следовательно, изучение аденилатциклазной системы необходимо для понимания патофизиологии и лечения многих болезней. Исследование роли вторичного медиатора циклического АМФ расширило наши знания об эндокринной, нервной и сердечнососудистой регуляции. И наоборот, исследования, ставившие своей целью разгадать биохимические основы определенных заболеваний, способствовали пониманию молекулярных механизмов, регулирующих синтез циклического АМФ.
Биохимия. Последовательность действия ферментов, участвующих в реализации эффектов гормонов (первичных медиаторов) осуществляющихся через циклический АМФ, представлена на рис. 67-1, а перечень гормонов, действующих с помощью этого механизма, приведен в табл. 67-1. Деятельность этих гормонов инициируется их связыванием со специфическими рецепторами, расположенными на наружной поверхности плазматической мембраны. Комплекс гормон — рецептор активирует связанный с мембраной фермент аденилатциклазу, которая синтезирует циклический АМФ из внутриклеточного АТФ. Внутри клетки циклический АМФ передает информацию от гормона, связываясь с собственным рецептором и активируя этот рецептор-зависимую от циклического АМФ протеинкиназу. Активированная протеинкиназа передает концевой фосфор АТФ специфическим белковым субстратам (как правило, ферментам). Фосфорилирование этих ферментов усиливает (или в некоторых случаях угнетает) их каталитическую активность. Измененная активность этих ферментов и вызывает характерное действие определенного гормона на его клетку-мишень.
Второй класс гормонов действует путем связывания с мембранными рецепторами, которые ингибируют аденилатциклазу. Действие этих гормонов, обозначаемых Ни, в отличие от стимулирующих гормонов (Не) описано ниже более детально. На рис. 67-1 показаны также дополнительные биохимические механизмы, ограничивающие действие циклического АМФ. Эти механизмы также могут регулироваться при участии гормонов. Это позволяет осуществлять тонкую настройку функции клеток с помощью дополнительных нервных и эндокринных механизмов.
Биологическая роль циклического АМФ. Каждая из белковых молекул, участвующих в сложных механизмах стимулирования — угнетения, представленных на рис. 67-1, представляет собой потенциальное место регуляции гормональной реакции на терапевтическое и токсическое действие лекарственных средств и на патологические изменения,, возникающие в ходе заболевания. Специфические примеры таких взаимодействий обсуждаются в последующих разделах этой главы. Для сведения их воедино следует рассмотреть общие биологические функции АМФ в качестве вторичного медиатора, что целесообразно сделать на примере регуляции процесса высвобождения глюкозы из запасов гликогена, содержащихся в печени (биохимическая система, в которой был обнаружен циклический АМФ), с помощью глюкагона и других гормонов.
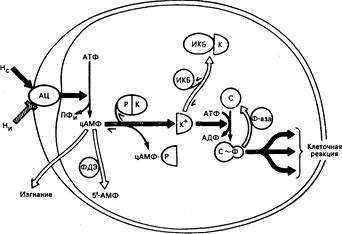
Рис. 67-1. Циклический АМФ — вторичный внутриклеточный медиатор для гормонов.
На рисунке изображена идеальная клетка, содержащая молекулы белка (ферменты), участвующие в медиаторных действиях гормонов, осуществляемых через циклический АМФ. Черные стрелки указывают путь потока информации от стимулирующего гормона (Не) до клеточной реакции, в то время как светлые стрелки указывают направление противоположных процессов, модулирующих или ингибирующих поток информации. Внеклеточные гормоны стимулируют (Не) или ингибируют (Ни) мембранный фермент — аденилатциклазу (АЦ) (см. описание в тексте и рис. 67-2). АЦ превращает АТФ в циклический АМФ (цАМФ) и пирофосфат (ПФи). Внутриклеточная концентрация циклического АМФ зависит от соотношения между скоростью его синтеза и характеристиками двух других процессов, направленных на выведение его из клетки: расщепление циклической нуклеотидной фосфодиэстеразой (ФДЭ), которая превращает циклический АМФ в 5'-АМФ, и удаление из клетки энергетически-зависимой транспортной системой. Внутриклеточные эффекты циклического АМФ опосредуются или регулируются белками по меньшей мере пяти дополнительных классов. Первый из них — зависимая от цАМФ протеинкиназа (ПК) — состоит из регулирующих (Р) и каталитических (К) субъединиц. В голоэнзиме ПК субъединица К каталитически неактивна (ингибирована субъединицей Р). Циклический АМФ действует, связываясь с субъединицами Р, высвобождая субъединицы К из комплекса цАМФ-Р. Свободные каталитические субъединицы (К+) катализируют передачу концевого фосфора АТФ в специфические белковые субстраты (С), например, фосфорилазкиназу. В фосфорилированном состоянии (С~Ф) эти белковые субстраты (обычно ферменты) инициируют характерные эффекты циклического АМФ внутри клетки (например, активацию гликогенфосфорилазы, ингибирование гликогенсинтетазы). Доля белковых субстратов киназы в фосфорилированном состоянии (С~Ф) регулируется белками двух дополнительных классов: ингибирующий киназу белок (ИКБ) обратимо связывается с К^, делая ее каталитически неактивной (ИКБ—К) Фосфатазы (Ф-аза) превращают С~Ф обратно в С, отнимая ковалентно связанный фосфор.
Перенос гормональных сигналов через плазматическую мембрану. Биологическая стабильность и структурная сложность пептидных гормонов, подобных глюкагону, делают их носителями разнообразных гормональных сигналов между клетками, но ослабляют их способность проникать через клеточные мембраны. Гормончувствительная аденилатциклаза позволяет информационному содержанию гормонального сигнала проникать через мембрану, хотя сам гормон не может проникнуть через нее.
Таблица 67-1. Гормоны, для которых циклический АМФ служит в качестве вторичного медиатора
| Гормон | Мишень:орган/ткань | Типичное действие |
| Адренокортикотропный гормон | Кора надпочечников | Продуцирование корти-зола |
| Кальцитонин | Кости | Концентрация кальция в сыворотке крови |
| Катехоламины (b-адре-нергические) | Сердце | Частота сердечных сокращений, сократимость миокарда |
| Хорионический гонадо-тропин | Яичники, семенники | Продуцирование половых гормонов |
| Фолликулостимулирую-щий гормон | Яичники, семенники | Гаметогенез |
| Глюкагон | Печень | Гликогенолиз, высвобождение глюкозы |
| Лютеинизирующий гормон | Яичники, семенники | \ Продуцирование половых гормонов |
| Рилизинг-фактор лютеи-низирующего гормона | Гипофиз | f Высвобождение лютеи-низирующего гормона |
| Меланоцитстимулирую-щий гормон | Кожа (меланоциты) | T Пигментация |
| Гормон паращитовидных желез | Кости, почки | T Концентрация кальция в сыворотке крови [ концентрация фосфора в сыворотке крови |
| Простациклин, проста-гландин е| | Тромбоциты | [ Агрегация тромбоцитов |
| Тиреотропный гормон | Щитовидная железа | T Продуцирование и высвобождение Тз и Т4 |
| Рилизинг-фактор тирео-тропного гормона | Гипофиз | f Высвобождение тирео-тропного гормона |
| Вазопрессин | Почки | f Концентрация мочи |
Примечание. Здесь перечислены только наиболее убедительно подтвержденные эффекты, опосредуемые циклическим АМФ, хотя многие из этих гормонов проявляют многочисленные действия в различных органах-мишенях.
Усиление. Связываясь с небольшим числом специфических рецепторов (вероятно, меньшим, чем 1000 на клетку), глюкагон стимулирует синтез гораздо большего числа молекул циклического АМФ. Эти молекулы в свою очередь стимулируют зависимую от циклического АМФ протеинкиназу, которая вызывает активацию тысяч молекул содержащейся в печени фосфорилазы (фермента, ограничивающего распад гликогена) и последующее высвобождение миллионов молекул глюкозы из единичной клетки.
Метаболическая координация на уровне единичной клетки. Помимо того что обусловленное циклическим АМФ фосфорилирование белка стимулирует фосфорилазу и способствует превращению гликогена в глюкозу, этот процесс одновременно дезактивирует фермент, синтезирующий гликоген (гликогенсинтетазу), и стимулирует ферменты, вызывающие глюконеогенез в печени. Таким образом, единичный химический сигнал — глюкагон — мобилизует энергетические резервы посредством нескольких путей метаболизма.
Преобразование разнообразных сигналов в единую метаболическую программу. Поскольку содержащаяся в печени аденилатциклаза может стимулироваться адреналином (действующим через b-адренорецепторы) так же, как и глюкагоном, циклический АМФ позволяет двум гормонам, обладающим различным химическим строением, регулировать углеводный обмен в печени. Если бы не существовало вторичного медиатора, то каждый из регулирующих ферментов, участвующих в мобилизации углеводов печени, должен был бы обладать способностью распознавать как глюкагон, так и адреналин.
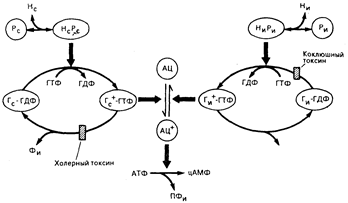
Рис. 67-2. Молекулярный механизм регуляции синтеза циклического АМФ гормонами, гормональными рецепторами и Г-белками. Аденилатциклаза (АЦ) в ее активной форме (АЦ+) превращает АТФ в циклический АМФ (цАМФ) и пирофосфат (ПФи). Активация и ингибирование АЦ опосредуются формально идентичными системами, показанными в левой и правой частях рисунка. В каждой из этих систем Г-белок колеблется между неактивным состоянием, будучи связанным с ГДФ (Г—ГДФ), и активным состоянием, будучи связанным с ГТФ (Г4"—ГТФ); только белки, находящиеся в активном состоянии, могут стимулировать (Гс) или ингибировать (Ги) активность АЦ. Каждый комплекс Г—ГТФ обладает внутренней активностью ГТФазы, которая превращает его в неактивный комплекс Г—ГДФ. Чтобы вернуть Г-белок в его активное состояние, стимулирующие или ингибирующие комплексы гормон—рецептор (НсРс и НиРи соответственно) способствуют замене ГДФ на ГТФ в месте связывания Г-белка с гуаниннуклеотидом. В то время как комплекс ГиР требуется для начальной стимуляции или ингибирования АЦ белками Гс или Гц, гормон может отсоединиться от рецептора независимо от регуляции АЦ, которая, напротив, зависит от длительности состояния связывания между ГТФ и соответствующим Г-белком, регулируемого его внутренней ГТФазой. Два бактериальных токсина регулируют активность аденилатциклазы, катализируя АДФ-рибозилирование Г-белков (см. текст). АДФ-рибозилирование Г с холерным токсином угнетает активность его ГТФазы, стабилизируя Гс в его активном состоянии и тем самым увеличивая синтез циклического АМФ. В противоположность этому АДФ-рибозилирование Ги коклюшным токсином предотвращает его взаимодействие с комплексом гнири и стабилизирует Ги в связанном с ГДФ неактивном состоянии; в результате этого коклюшный токсин предотвращает гормональное угнетение АЦ.
Координированная регуляция различных клеток и тканей первичным медиатором. В случае классической реакции на стресс «сражайся или беги» катехоламины связываются с b-адренорецепторами, расположенными в сердце, жировой ткани, кровеносных сосудах и многих других тканях и органах, включая печень. Если бы циклический АМФ не опосредовал большинство реакций на действие b-адренергических катехоламинов (например, увеличение частоты сердечных сокращений и сократимости миокарда, расширение сосудов, снабжающих кровью скелетную мускулатуру, мобилизация энергии из запасов углеводов и жиров), то совокупность огромного количества отдельных ферментов в тканях должна была бы обладать специфическими местами связывания для регуляции катехоламинами.
Аналогичные примеры биологических функций циклического АМФ можно было бы привести и в отношении других первичных медиаторов, приведенных в табл. 67-1. Циклический АМФ действует как внутриклеточный медиатор для каждого из этих гормонов, обозначая их присутствие на поверхности клетки. Подобно всем эффективным медиаторам, циклический АМФ обеспечивает простой, экономичный и высокоспециализированный путь передачи разнородных и сложных сигналов.
Гормончувствительная аденилатциклаза. Основным ферментом, опосредующим соответствующие эффекты этой системе, является Гормончувствительная аденилатциклаза. Этот фермент состоит по меньшей мере из пяти классов разделимых белков, каждый из которых внедрен в жировую двухслойную плазматическую мембрану (рис. 67-2).
На наружной поверхности клеточной мембраны обнаруживаются два класса гормональных рецепторов, Рс и Ри. Они содержат специфические участки распознавания для связывания гормонов, стимулирующих (Нс) или ингибирующих (Ни) аденилатциклазу.
Каталитический элемент аденилатциклазы (АЦ), обнаруживаемый на цитоплазматической поверхности плазменной мембраны, превращает внутриклеточный АТФ в циклический АМФ и пирофосфат. На цитоплазматической поверхности присутствуют также два класса гуаниннуклеотидсвязывающих регулирующих белков. Эти белки, Гс и Ги, опосредуют стимулирующее и ингибирующее действие, воспринимаемое рецепторами Рс и Ри соответственно.
Как стимулирующая, так и угнетающая парные функции белков зависят от их способности связывать гуанозинтрифосфат (ГТФ) (см. рис. 67-2). Только ГТФ-связанные формы Г-белков регулируют синтез циклического АМФ. Ни стимуляция, ни угнетение АЦ не являются постоянным процессом; вместо этого концевой фосфор ГТФ в каждом комплексе Г—ГТФ в конце концов гидролизируется, а Гс—ГДФ или Ги — ГДФ не могут регулировать АЦ. По этой причине стойкие процессы стимуляции или угнетения аденилатциклазы требуют непрерывного превращения Г—ГДФ в Г—ГТФ. В обоих проводящих путях комплексы гормон — рецептор (НсРс или НиРи) усиливают превращение ГДФ в ГТФ. Этот рециркуляционный во временном и пространственном отношениях процесс отделяет связывание гормонов с рецепторами от регуляции синтеза циклического АМФ, используя энергетические запасы в концевой фосфорной связи ГТФ для усиления действия комплексов гормон — рецептор.
Эта схема объясняет, каким образом несколько разных гормонов могут стимулировать или угнетать синтез циклического АМФ в пределах единичной клетки. Поскольку рецепторы по своим физическим характеристикам отличаются от аденилатциклазы, совокупность рецепторов, находящихся на поверхности клетки, определяет специфическую картину ее чувствительности к внешним химическим сигналам. Отдельная клетка может иметь три или более различных рецептора, воспринимающих угнетающее действие, и шесть или более отличающихся от них рецепторов, воспринимающих стимулирующее действие. И напротив, все клетки, по-видимому, содержат сходные (возможно, идентичные) компоненты Г и АЦ.
Молекулярные компоненты гормончувствительной аденилатциклазы обеспечивают контрольные точки для изменения чувствительности данной ткани к гормональной стимуляции. Как Р, так и Г-компоненты служат решающими факторами физиологической регуляции чувствительности к гормонам, и изменения Г-белков рассматривают как первичное поражение, возникающее при четырех обсуждаемых ниже заболеваниях.
Регуляция чувствительности к гормонам (см. также гл. 66). Повторное введение какого-либо гормона или лекарственного средства, как правило, вызывает постепенное повышение резистентности к их действию. Этот феномен носит разные названия: гипосенсибилизация, рефрактерность, тахифилаксия или толерантность.
Гормоны или медиаторы могут вызвать развитие гипосенсибилизации, являющейся рецепторспецифичной, или «гомологичной». Например, введение b-адренергических катехоламинов вызывает специфическую рефрактерность миокарда к повторному введению этих аминов, но не к тем лекарственным средствам, которые не действуют через b-адренорецепторы. Рецепторспецифическая гипосенсибилизация включает в себя по меньшей мере два отдельных механизма. Первый из них, быстро развивающийся (в течение нескольких минут) и быстро обратимый при удалении введенного гормона, функционально «расцепляет» рецепторы и Гс-белок и, следовательно, снижает их способность стимулировать аденилатциклазу. Второй процесс связан с фактическим уменьшением числа рецепторов на клеточной мембране — процесс, называемый рецептороуменьшающей регуляцией. Процесс рецептороуменьшающей регуляции для своего развития требует несколько часов и является труднообратимым.
Процессы гипосенсибилизации представляют собой часть нормальной регуляции. Устранение нормальных физиологических стимулов может привести к повышению чувствительности ткани-мишени к фармакологической стимуляции, как это происходит при развитии гиперчувствительности, вызванной денервацией. Потенциально важная клиническая корреляция такого увеличения числа рецепторов может развиться у больных при внезапном прекращении лечения анаприлином, являющимся b-адреноблокирующим средством. У таких больных часто наблюдаются преходящие признаки повышенного симпатического тонуса (тахикардия, повышение артериального давления, головные боли, дрожание и т. д.) и могут развиться симптомы коронарной недостаточности. В лейкоцитах периферической крови больных, получающих анаприлин, обнаруживают повышенное число b-адренорецепторов, и число этих рецепторов медленно возвращается к нормальным значениям при прекращении приема препарата. Хотя более многочисленные другие рецепторы лейкоцитов не опосредуют сердечно-сосудистые симптомы и явления, возникающие в случае отмены анаприлина, рецепторы в миокарде и других тканях, вероятно, претерпевают такие же изменения.
Чувствительность клеток и тканей к гормонам может регулироваться и «гетерологичным» путем, т. е. когда чувствительность к одному гормону регулируется другим гормоном, действующим через иной набор рецепторов. Регуляция чувствительности сердечно-сосудистой системы к b-адренергическим аминам гормонами щитовидной железы является самым известным клиническим примером гетерологичной регуляции. Гормоны щитовидной железы вызывают накопление избыточного количества b-адренорецепторов в миокарде. Это увеличение. числа рецепторов частично объясняет повышенную чувствительность сердца больных гипертиреозом к катехоламинам. Однако тот факт, что у экспериментальных животных увеличение числа b-адренорецепторов, вызываемое введением гормонов щитовидной железы, недостаточно для того, чтобы отнести на его счет повышение чувствительности сердца к катехоламинам, позволяет предположить, что влиянию гормонов щитовидной железы подвержены также и компоненты реакции на гормоны, действующие дистальнее рецепторов, возможно включающие в себя Гс, но не ограничивающиеся этими субъединицами. К числу других примеров гетерологичной регуляции относятся контролирование эстрогеном и прогестероном чувствительности матки к расслабляющему действию b-адренергических агонистов и повышенная реактивность многих тканей по отношению к адреналину, вызываемая глюкокортикоидами.
Второй тип гетерологичной регуляции заключается в угнетении гормональной стимуляции аденилатциклазы веществами, действующими через Ри и Ги, как отмечалось выше. Ацетилхолин, опиаты и a-адренергические катехоламины действуют через отличные друг от друга классы воспринимающих ингибирующее действие рецепторов (мускариновые, опиатные и a-адренорецепторы), снижая чувствительность аденилатциклазы определенных тканей к стимулирующему действию других гормонов. Хотя клиническое значение гетерологичной регуляции этого типа не установлено, угнетение синтеза циклического АМФ морфином и другими опиатами могло бы быть причиной некоторых аспектов толерантности к препаратам этого класса. Аналогично устранение такого угнетения может играть определенную роль в развитии синдрома, следующего за прекращением введения опиатов.
Псевдогипопаратиреоз (см. также гл. 336). Генетический дефект, затрагивающий Гс-компонент аденилатциклазы, служит молекулярной основой для развития псевдогипопаратиреоза типа 1 (ПГП-1). Это редкое наследственное заболевание характеризуется гипокальциемией и гиперфосфатемией, повышенным уровнем содержания гормона паращитовидных желез в сыворотке крови (ГПЖ) и резистентностью к метаболическим эффектам вводимого экзогенного ГПЖ.
ГПЖ регулирует гомеостаз кальция, по меньшей мере частично, посредством стимуляции аденилатциклазы в почках и костях. Введение ГПЖ здоровым людям (и больным, страдающим идиопатическим или развившимся в результате хирургического вмешательства гипопаратиреозом) вызывает резкое увеличение экскреции циклического АМФ в мочу. В противоположность этому введение ГПЖ вызывает лишь небольшое увеличение (или вообще не изменяет) концентрации циклического АМФ в моче у больных, страдающих ПГП-1. Первоначально обнаружение этого факта породило предположение о том. что причиной ПГП-1 служит дефект в рецепторах ГПЖ, снижающий их способность стимулировать синтез циклического АМФ.
Однако влиянием дефекта, ограниченного только рецепторами ГПЖ, нельзя объяснить тот факт, что многие страдающие ПГП-1 больные также частично резистентны к действию других гормонов, включая тнреотропный (ТТГ), антидиуретический гормоны, глюкагон и гонадотропины. Действительно, некоторые страдающие ПГП-1 больные нуждаются в заместительной терапии по поводу симптоматического гипотиреоза вследствие резистентности щитовидной железы к действию ТТГ. Резистентность к действию других гормонов по большей части клинически не проявляется и ее можно выявить только с помощью специальных тестов.
То, что циклический АМФ служит для этих гормонов в качестве вторичного медиатора, позволяет предположить, что причиной ПГП-1 является дефект, действующий дистальнее гормональных рецепторов и изменяющий некий компонент, общий для всех опосредуемых циклическим АМФ реакций. Для многих людей, страдающих ПГП-1, такой компонент—Гс-белок, активность которого снижена в эритроцитах, тромбоцитах и фибробластах кожи у большинства больных приблизительно на 50%. У одного из больных было выявлено снижение активности Гс в почках, и, вероятно, аналогичные процессы наблюдались и в других клетках, являющихся эндокринными мишенями, включая кости, щитовидную железу, печень и т. д.
Возникает вопрос, если дефектность Гс — всеобщее явление при ПГП-1, то почему большинство из важных клинических последствий этого связано с резистентностью к единственному гормону — ГПЖ? Хотя на этот вопрос нельзя ответить определенно, вероятно, что большинство опосредуемых циклическим АМФ реакций на действие гормонов поддерживаются, несмотря на частичную неполноценность Гс, рядом компенсаторных механизмов, включая увеличение концентраций циркулирующих в крови стимулирующих гормонов. Однако наличие повышенной концентрации циркулирующего в крови ГПЖ оказывается недостаточным фактором для поддержания нормального гомеостаза кальция при ПГП-1, предположительно вследствие того, что нормальное функционирование ГПЖ решающим образом определяется условием нормальной активности Гс. К счастью, нарушения реактивности ГПЖ можно ликвидировать лечением витамином D, с помощью которого концентрации кальция и фосфора в сыворотке крови восстанавливаются до нормы.
Холера (см. также гл. 115). Повышение содержания циклического АМФ в клетках слизистой оболочки кишечника вызывает массивную секрецию воды и электролитов, что приводит к развитию холерной диареи. Патогенный Vibrio cholerae продуцирует белковый экзотоксин, способный стимулировать синтез циклического АМФ фактически во всех клетках организма. Клинически заболевание ограничивается слизистой оболочкой кишечника, поскольку токсин не абсорбируется из пищеварительного тракта, таким образом, другие ткани недоступны для токсина, вырабатываемого бактериями, находящимися в кишечнике.
В отличие от процесса стимуляции аденилатциклазы гормонами действие холерного токсина начинается постепенно и не прекращается немедленно после введения токсина. Причиной такого различия является то, что холерный токсин действует не посредством обратимого связывания со стимулирующим рецептором, а как фермент, образуя стабильную ковалентную модификацию компонента Гс-белка аденилатциклазы. После связывания токсина с клетками одна из его пептидных субъединиц проникает через клеточную мембрану, где катализирует АДФ-рибозилирование Гс, используя внутриклеточный никотинамидадениндинуклеотид (НАД ) в качестве субстрата:
Гс + НАД+ ---токсин---> Гс-АДФ-рибоза + никотинамид + Н+.
АДФ-рибозилирование Гс увеличивает синтез циклического АМФ, по-видимому, путем снижения скорости гидролиза ГТФ в комплексе Гс—ГТФ-С, который синтезирует циклический АМФ (см. рис. 67-2).
Этот биохимический механизм помогает понять причину развития холерной диареи в результате жизнедеятельности относительно небольшого числа патогенных микроорганизмов, а также то, почему болезнь может продолжаться некоторое время и после ликвидации микроорганизмов из кишечника. Первый феномен объясняется тем, что данный токсин является ферментом и поэтому небольшого числа его молекул достаточно для АДФ-рибозилирования значительной части молекул Гс в клетке. Сохраняющийся повышенным уровень синтеза циклического АМФ после выведения токсина (по меньшей мере в экспериментальных исследованиях) коррелирует со стабильностью Гс-АДФ-рибозы. Большинство клеток, по-видимому, 'не содержат ферментов, способных отделить АДФ-рибозу от Гс, и поэтому действие токсина прекратится только тогда, когда молекулы Гс-АДФ-рибозы будут замещены заново синтезированными молекулами Гс. Для выздоровления может потребоваться даже замена самих клеток слизистой оболочки. Выяснение молекулярной основы действия холерного токсина послужило ценным инструментом обнаружения Гс и определения его характеристик, а также повысило уровень нашего знания биохимической основы действия гормонов.
Коклюш (см. также гл. 109). Молекулярный патогенез коклюша в. бронхах очень близок к патогенезу холеры в кишечнике. Ни один из этих двух микроорганизмов не внедряется в ткани, —оба они вызывают заболевание, вырабатывая экзотоксины, изменяющие синтез циклического АМФ в клетках хозяина. Bordetella pertussis секретирует два патогенных экзотоксина: молекулярной мишенью одного из них, называемого коклюшевым токсином, является связанный с гуаниннуклеотидом угнетающий белок аденилатциклазы. Другой токсин сам является молекулой аденилатциклазы.
Подобно холерному, коклюшевый токсин катализирует перенос АДФ-рибозы от НАД+ к белку мембраны; в данном случае это белок Ги. АДФ-рибозилированная форма Ги не способна взаимодействовать с Ри (см. рис. 67-2); в результате этого коклюшный токсин предотвращает угнетение аденилатциклазы ингибирующими лигандами, такими как мускариновые, a-адренергические и опиатные агонисты.
Ключевые бронхиальные клетки-мишени для коклюшного токсина и молекулярные события, связывающие АДФ-рибозилирование Г„ с симптомами коклюша, в значительной степени неизвестны. Однако реакция АДФ-рибозилирования помогает объяснить одно загадочное явление: лечение коклюшной инфекции антибиотиками в очень незначительной степени уменьшает тяжесть и длительность заболевания; и действительно, симптомы болезни могут сохраняться в течение трех и более недель после исчезновения микроорганизмов из трахеобронхиального дерева. АДФ-рибозилирование Ги (подобно АДФ-рибозилированию Гс, описанному выше) трудно обратимо с помощью клеточных ферментов. Вероятно, болезнь не отступит даже в отсутствие поступления дополнительных порций токсина, пока не будут замещены модифицированные белки Ги или клетки, содержащие такие белки.
Второй секретируемый В. pertussis патогенный продукт — аденилатциклазный токсин — остается ферментно инертным до тех пор, пока не внедрится в клетки хозяина. Там он активируется путем связывания с кальмодулином, в результате чего происходит существенное увеличение содержания циклического АМФ в клетке. В отличие от коклюшевого действие аденилатциклазного токсина быстро исчезает после удаления его источника; клеточные ферменты разрушают бактериальный фермент и концентрация циклического АМФ в клетке быстро возвращается к норме.
Хотя большинство связей между молекулярными действиями этих токсинов и развивающимся в результате этого заболеванием недостаточно ясны, эксперименты in vitro показывают, что оба токсина могут нарушать функцию нейтрофильных гранулоцитов человека—клеток, играющих основную роль в защите организма от инфекций. Накопление циклического АМФ, вызываемое аденилатциклазным токсином, нарушает способность нейтрофильных гранулоцитов убивать захваченные микроорганизмы. Коклюшный токсин путем АДФ-рибозилирования Ги или Ги-подобных молекул блокирует реакции нейтрофильных гранулоцитов (хемотаксис, высвобождение лизосомальных гидролаз и т. д.) на комплемент и другие факторы хемотоксина. Любой из этих двух эффектов или оба они могут способствовать увеличению чувствительности к легочным инфекциям, вызываемым другими микроорганизмами, что является частым осложнением коклюша.
Помимо их роли в качестве инструментов для исследования преобразования гормональных сигналов, знание этих токсинов может принести реальную пользу в практической медицине. Хотя современная практика иммунизации экстрактами целых коклюшевых бактерий эффективно предотвращает инфицирование, сама по себе иммунизация является причиной значительного числа случаев заболевания. Возбудители коклюша, специфически неспособные продуцировать любой из двух описанных выше токсинов, делаются безвредными для организма хозяина. В соответствии с этим иммунизация препаратами очищенного токсина должна предотвращать развитие болезни и снизить заболеваемость, вызываемую самой иммунизацией. Результаты предварительных клинических испытаний подкрепляют оба эти прогноза.
Сибирская язва (см. также гл. 98). Кожное инфицирование Bacillus antracis вызывает развитие характерного поражения с областью центрального некроза, окруженной выступающим подкожным отеком. В то время как некротические поражения, по-видимому, не связаны с участием системы циклического АМФ, «отечным фактором» в организме является аденилатциклаза. Подобно аденилатциклазному токсину коклюша отечный фактор получает доступ к клеткам хозяина, активируется клеточным кальмодулином и увеличивает внутриклеточную концентрацию циклического АМФ. Интересно отметить, что встречаются редкие случаи, когда у больных, в организм которых с пищей попали В. antracis, развилась водянистая диарея, неотличимая от такого же симптома холеры. Диарея в таких случаях, вероятно, является результатом проникновения отечного фактора внутрь клеток слизистой оболочки кишечника, приводящего к увеличению концентрации циклического АМФ и секреции соли и воды.
Циклический АМФ в клинической медицине. Большое число гормонов и медиаторов действуют путем стимуляции аденилатциклазы, а некоторые фармакологические антагонисты действуют путем блокирования их связывания со специфическими рецепторами — например, анаприлина с b-адренорецепторами и циметидина с На-гистаминовыми рецепторами. Терапевтическое действие этих веществ зависит от увеличения или уменьшения концентрации циклического АМФ в клетках-мишенях и тканях больных. Кроме этого, метилксантины (кофеин и теофиллин) блокируют фосфодиэстеразы циклических нуклеотидов и могут оказывать некоторые из своих лечебных воздействий (например, расширение бронхов) путем увеличения концентрации циклического АМФ в клетках.
В клинической практике определение содержания циклического АМФ в моче целесообразно при диагностике заболеваний, сопровождающихся нарушением гомеостаза кальция и концентрации ГПЖ. Значительная часть присутствующего в моче циклического АМФ образуется в клетках проксимальных канальцев, реагирующих на циркулирующий в крови ГПЖ. Таким образом, уровень содержания циклического АМФ в моче обеспечивает удобное «окно», заглянув в которое можно оценить действие ГПЖ на Почки и которое может отражать повышение концентрации ГПЖ (при гиперпаратиреозе), снижение ее (при гипопаратиреозе) или резистентность конечного органа к действию ГПЖ (при ПГП-1) (см. гл. 336).
Однако в настоящее время реальное значение циклического АМФ для медицины заключается в том, что он является инструментом для исследования и понимания нормальной и патологической регуляции, а также для разработки новых лекарственных средств. Образцы аденилатциклазы в настоящее время обычно используют для проверки новых веществ на их способность стимулировать или блокировать адренергические, гистаминергические и многие пептидные рецепторы. Гормональные рецепторы не являются единственными основными и специфическими точками контроля регуляции, опосредуемой через циклический АМФ; вероятно, что и другие белки (см. рис. 67-1) будут служить мишенями для новых лекарственных средств в будущем.
Прочие вторичные медиаторы. Хотя наиболее хорошо изученным гормональным вторичным медиатором является циклический АМФ, некоторые гормоны действуют посредством увеличения внутриклеточных концентраций других химических сигнальных веществ, включая ионы кальция и циклической гуанозин-3',5^монофосфат (циклический ГМФ). Например, определенные эффекты a-адренергических и холинергических (мускариновых) веществ, по-видимому, опосредуются повышенными концентрациями кальция в цитоплазме. Клетки многих типов содержат гуанилатциклазу, фосфодиэстеразы циклического ГМФ и белковые киназы, которые специфически стимулируются циклическим ГМФ. Тем не менее роль этого вторичного циклического нуклеотида в нормальной и патологической регуляции не совсем ясна.
Дата добавления: 2016-03-05; просмотров: 1399;