Живые организмы как среда жизни. В течение всей жизни или части жизненного цикла многие виды гетеротрофных организмов обитают в других живых организмах
В течение всей жизни или части жизненного цикла многие виды гетеротрофных организмов обитают в других живых организмах, тела которых служат для них средой, значительно отличающейся от внешней по своим свойствам. Использование одними живыми организмами других в качестве среды обитания —древнее и широко распространенное явление в природе. Установлено, что прокариотические организмы (бактерии, актиномиценты, сине-зеленые водоросли) имеют сожителей. У большого числа одноклеточных эукариотических форм (красные, зеленые и диатомо-вые водоросли, амебы, радиолярии и др.) обнаружены внутриклеточные паразиты и симбионты. Практически нет ни одного вида многоклеточных организмов, не имеющих внутренних обитателей. Чем выше организация хозяев, чем больше степень дифференцированнос-ти их тканей и органов, тем более разнообразные условия они могут предоставить своим сожителям. Так, английский ученый А. Е, Шит-ли писал, что каждая птица — это, по сути, настоящий летающий зоопарк. Разнообразие мельчайших существ, живущих на птицах, поистине ошеломляюще (рис. 5.46).
Перья служат пищей вшам и клещам; кожей питаются некоторые мухи; блохи, вши, москиты, пиявки и другие паразиты сосут кровь птиц, находясь на поверхности тела, в то время как представители простейших разрушают красные кровяные тельца внутри организма. Практически в любых органах птицы можно обнаружить разнообразных паразитических червей.

Рис. 5.46. Живые организмы как среда жизни
(на примере паразитов птицы), по П. Фарбу, 1971
Джонатан Свифт также обратил внимание на огромную распространенность явления паразитизма:
Под микроскопом он открыл, что на блохе
живет блоху кусающая блошка;
На блошке той — блошинка-крошка,
В блошинку же вонзает зуб сердито
Блошиночка... и так ad infinitum.
Паразитизм — явление столь всеобщее, что единственные живые существа, не подверженные нападению паразитов, это те паразиты, которые представляют собой последнее звено длинной цепи питания. Однако чем ниже на эволюционной лестнице находится та или иная группа живых организмов, тем больше она включает видов паразитов. Некоторые группы низших животных, особенно это касается плоских червей, нематод и некоторых членистоногих, состоят исключительно из паразитических форм. У позвоночных паразитизм как способ существования встречается крайне редко. В царстве растений паразиты широко распространены среди грибов (рис. 5.47).

Рис. 5.47. Картофель, пораженный фитофторой
(возбудитель — низший гриб Phytophthora infestans)
Несколько паразитирующих видов есть и среди высших цветковых растений, таких, как омела, повилика и др. Классический пример — виды рода (Rafflesia), у которых вегетативное тело — это нити, напоминающие гифы гриба, погруженные в ткани питающего растения. Снаружи развиваются лишь огромные (до 1 м в диаметре) цветки (рис. 5.48).
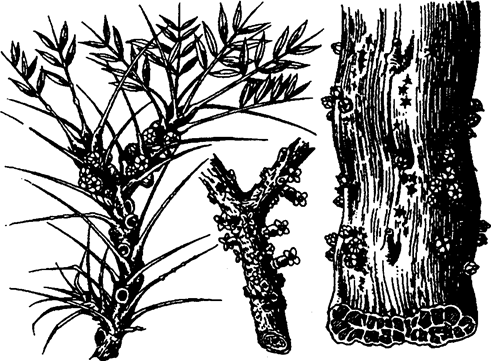
Рис. 5.48. Растение из семейства Rafflesiaceae —
Эндопаразита древесных видов (по А. Кернеру, 1896):
тело паразита находится внутри ветви или ствола хозяина, снаружи видны лишь цветки или плоды
Для животных и растений, ведущих паразитический образ жизни, организм, на котором или в котором они поселяются (хозяин), является специфической средой обитания. Большая часть паразитов практически полностью утратила связь с внешним миром, и все стадии их развития происходят в организме хозяев, например малярийный плазмодий и др.
Между паразитами и хозяевами в процессе эволюции возникли сложные взаимоотношения. Различные их оттенки отражают пути возникновения паразитизма.
Первый путь — простое «квартирантство», как на рис.5.49 (2).

Рис. 5.49. Животное-паразитоид наездник, откладывающий яйца в тлю (1); «квартирантство». Самка горчака откладывает икру в мантийную полость двустворчатого моллюска беззубки (2)
Нередко более мелкий организм поселяется в жилище более крупного или вблизи него и со временем переходит на тело хозяина, а затем и внутрь, переключаясь на питание за счет его жизни или соков и таким образом причиняя ему вред. Квартирант может превратиться в конечном итоге в паразита, а тело хозяина становится для него средой обитания.
Второй путь перехода к паразитизму — через хищничество. Так, хищник при нападении на крупную добычу, которую не может уничтожить и съесть сразу, при определенных условиях проникнув внутрь тела хозяина и найдя там благоприятную среду — обилие пищи, может превратиться в паразита. Организм хозяина для паразита становится средой обитания.
Третий путь — случайное проникновение будущего паразита в организм хозяина. Например, крупные животные могут заглатывать с пищей мелкие формы, некоторые из них не погибают, а, приспосабливаясь к новым условиям, превращаются в паразитов. В природе сохранилось немало примеров перехода этих трех путей от квартирантства, хищничества и случайного паразитиро-вания к подлинному паразитизму. Вместе с тем сегодня трудно сказать, когда на Земле появился первый паразит, от какой группы животных или растений он произошел.
Паразитов обычно делят на две группы: эктопаразитов и эндопаразитов. Эктопаразиты — это наружные паразиты, обитающие на поверхности тела хозяина (клещи, пиявки, блохи). У растений-эктопаразитов большая часть тела находится вне хозяина, а в него внедряются и вступают в контакт с живыми клетками лишь органы чужеядного питания — присоски или гаустории (повилика европейская — Cuscuta europaea и др.). Эндопаразиты — внутренние паразиты, живущие внутри тела хозяина. Это большинство гельминтов, бактерии, вирусы, паразитические простейшие. У растений-эндопаразитов почти все тело помещается внутри тканей хозяина, наружу выходят лишь органы размножения (виды рода Rafflesia, см. рис. 5.48). У многих паразитических грибов тело находится в межклетниках высшего растения, а в клетки внедряются гаустории. Паразитические низшие грибы и бактерии живут внутри клеток растения-хозяина.
Отличают и стационарный паразитизм, когда паразит на длительное время, часто на всю жизнь, связывает себя с хозяином. Стационарные паразиты могут быть приурочены к одному хозяину (постоянные)— вши, пухоеды, чесоточные зудни, или развитие их протекает со сменой хозяев (периодические) — многие ленточные черви, сосальщики. Так, малярийный плазмодий определенную часть жизни проводит в малярийном комаре — окончательный хозяин. Промежуточным хозяином является человек. Окончательным хозяином служит тот организм, в котором обитает половозрелая форма паразита, а промежуточным — в котором паразит проходит личиночную, неполовозрелую стадию.
Клещевой энцефалит—заболевание, поражающее центральную нервную систему человека. Оно вызывается вирусом, переносчики и хранители вируса— иксодовые клещи. Излюбленные места обитания клещей — южная часть таежных лесов на всем протяжении европейской и азиатской частей России.
Имеются и временные паразиты. Они не всю свою жизнь связывают с хозяином, а часть ее проводят свободно. К ним относят кровососущих двукрылых и многих клопов.
Таким образом, еще раз обращаем внимание, что паразитов больше всего среди микроорганизмов и сравнительно примитивных многоклеточных, а подверженность паразитизму наиболее развита у позвоночных животных и цветковых растений.
Паразиты обитают в специфических условиях внутренней среды хозяина. С одной стороны, это дает им ряд экологических преимуществ, а с другой — затрудняет осуществление их жизненного цикла по сравнению со свободноживущими видами.
Одним из важных преимуществ паразитов является обильное их снабжение пищей за счет содержимого клеток, соков и тканей тела хозяина или содержимого кишечника. Обильная и легкодоступная пища служит условием быстрого роста паразитов. Так, в кишечном тракте позвоночных паразиты достигают больших размеров по сравнению с их свободноживущими родственниками.
Человеческая и свиная аскариды — наиболее крупные представители класса нематод, а бычий и свиной солитеры, лентец широкий — гиганты среди плоских червей, достигающие в длину 8 — 12м, тогда как самые крупные тропические турбеллярии не превышают 60 см. Большинство свободноживущих инфузорий имеют размеры 50 — 100 мкм, тогда как сожители жвачных Entodinomorpha достигают 200 — 500 мкм, а в некоторых случаях 2 — 3мм, как Pycnothrix из кишечника даманов. Вторым важным экологическим преимуществом для обитателей живых организмов является их защищенность от непосредственного воздействия факторов внешней среды. Внутри хозяина они практически не встречаются с опасностью высыхания, резким колебанием температур, значительными изменениями солевого и осмотического режимов и т. п. Так, в особенно стабильных условиях существуют внутренние обитатели гомойотермных животных. Колебания условий внешней среды сказываются на внутренних паразитах и симбионтах лишь опосредованно, через организм хозяев. Практически неограниченные пищевые ресурсы служат для паразитов также условием высокого потенциала их размножения, которое обеспечивает им вероятность заражения других хозяев.
Защищенность от внешних врагов, обилие легкоусвояемой пищи, относительная стабильность условий делают ненужной сложную дифференцировку тела, отсюда многие внутренние паразиты и симбионты характеризуются в эволюции вторичным упрощением строения, включая потерю целых систем органов. Например, ленточные черви, всасывающие переваренную хозяином пищу через покровы, отличаются отсутствием пищеварительной системы и редукцией нервной. Галловые клещи, живущие в тканях растений, проводят всю свою жизнь и даже размножаются на стадии эмбриона всего с одной парой конечностей вместо четырех пар, как свойственно всем паукообразным.
В связи с паразитическим образом жизни у растений редуцируется ряд физиологических функций и соответствующих органов. Например, отсутствуют или сильно редуцированы корни. Потеря способности к фотосинтезу привела к отсутствию хлорофилла. Так, у некоторых заразих удается обнаружить лишь его следы. По мере усиления паразитических свойств в эволюционном ряду растений-паразитов сокращается ферментный аппарат. Остаются лишь специализированные ферменты, позволяющие паразитировать на узком круге хозяев. Эта биохимическая специализация послужила основой строгой избирательности многих паразитов по отношению к растениям-хозяевам.
Одним из наиболее известных эктопаразитов, как уже было отмечено выше, является повилика европейская (Cuscuta europaea), паразитирующая на многих травянистых видах и невысоких кустарниках (рис. 5.50).

Рис. 5.50. Повилика и заразиха:
1 — повилика клеверная; 2 — заразиха подсолнечниковая
Тело растения представлено тонкими желтоватыми стеблями, напоминающими нити, которые обвиваются вокруг стеблей растения-хозяина, нередко переплетаясь друг с другом. Внедрение в ткани хозяина происходит с помощью гаусторий. Листья у повилик полностью отсутствуют, а после прикрепления к хозяину отмирает и слабо развитый корень, поэтому стебли повилики не имеют связи с почвой. В середине лета на них появляются шаровидные клубочки мелких бледно-розовых цветков. Из других бес-хлорофилльных паразитов следует назвать виды рода заразихи (Orobanche), которые поражают многие сельскохозяйственные культуры (конопля, подсолнечник, табак и др.) и виды дикой флоры. Толстый мясистый стебель заразихи, покрытый чешуйчатыми бесцветными листьями, несет на конце колосовидное соцветие. Нижний его конец, обычно вздутый и утолщенный, прикреплен к корню растения-хозяина.
Существует ряд переходов от настоящих паразитов к автотроф-ным растениям. Промежуточное звено в нем представляют полупаразиты, которых нередко называют «зелеными паразитами». Это растения, частично или полностью утратившие способность поглощать из почвы воду и питательные вещества, но сохранившие хлорофилл и возможность самостоятельного фотосинтеза. Широко известны луговые травянистые полупаразиты. Среди них погремки большой и малый — Alectorolophus major и A. minor, мытник — Pedicularis, очанки — Euphrosia и др. К корням травянистых растений они прикрепляются с помощью гаусториев. Полупаразитный образ жизни ведут омела белая (Viscum album) и ремнецветник европейский (Loranthus europaeum), поселяющиеся на ветвях древесных пород, таких, как липа, тополь и др. Всего насчитывают 1964 вида полупаразитов.
Для паразитов выход во внешнюю среду, как правило, чреват многими опасностями, отсюда на той стадии жизненного цикла, которую паразиты проводят вне хозяина, у них развиваются различные защитные приспособления, позволяющие пережить этот критический период (толстые и многослойные оболочки яиц гельминтов, цисты кишечных амеб, ооцисты со спорами кокцидий, способность к анабиозу у ряда личинок нематод и т. д.). При отсутствии же в жизненном цикле паразитов стадии выхода во внешнюю среду, как у малярийного плазмодия, таких защитных приспособлений не обнаруживается.
Основные экологические трудности, с которыми сталкиваются внутренние сожители живых организмов, — это ограниченность жизненного пространства для внутриклеточных и тканевых обитателей, сложности снабжения кислородом, трудность распространения от одной особи хозяев к другим, а также защитные реакции организма хозяина против паразитов. Ограниченность жизненного пространства особенно сказывается на размерах и форме внутриклеточных паразитов. Так, грегарины, живущие в полости кишечника, — крупные споровики со сложно расчлененной клеткой, тогда как малярийные плазмоиды, являющиеся внутриклеточными паразитами, отличаются мелкими размерами и упрощенным внешним строением.
Недостаток кислорода в тканях и особенно в желудочно-ки-шечном тракте организмов-хозяев приводит к тому, что у многоклеточных обитателей внутриорганизменной среды вырабатывается преимущественно анаэробный тип обмена. Необходимая для работы клеток энергия высвобождается за счет разных видов брожения, а не за счет дыхания. Так, у человеческой аскариды утрачены все ферменты дыхательного цикла, кислород действует на них как яд, что и используется в медицинской практике. Однако целый ряд паразитов не утрачивает полностью способности к дыханию и может переключаться с анаэробного типа обмена на аэробный, таковы жгутиковые Trichomonas, эхинококк и др.
Среда обитания паразитов ограничена как во времени (жизнью хозяина), так и в пространстве. Поэтому основные адаптации направлены на возможность распространения в этой среде, передачи от одного хозяина к другому. Так, для паразита растений очень важно обеспечить контакт с хозяином, начиная с прорастания семян. Семена многих паразитических видов не прорастают в почве до тех пор, пока не окажутся вблизи корней растений-хозяев, от которых в почву поступают выделения, стимулирующие прорастание семян паразита и определяющие направления роста его гаусторий. Проростки некоторых паразитов (повилик, заразих) производят винтообразные движения «в поисках» корня или стебля растения-хозяина.
С помощью электронно-микроскопических исследований установлено, что вблизи проникновения гаустории омелы в ткани тополя направление роста сосудов растения-хозяина отклоняется в сторону присоски под влиянием выделений паразита, облегчающих установление контактов с хозяином.
Важная адаптация паразитов — синхронизация их жизненных циклов с сезонным развитием растений-хозяев, позволяющая осуществить заражение хозяина в нужный момент. Это явление широко известно для многих паразитических грибов. Некоторые паразиты способны к временной приостановке развития, начинающегося в «неподходящий» момент. Например, слишком рано появившиеся проростки повилики прекращают рост, иногда на несколько недель, до тех пор, пока рядом не разовьются проростки травянистых растений — возможных хозяев. Существенными адаптациями паразитов являются повышенная способность к размножению, выработка сложных жизненных циклов, использование переносчиков и промежуточных хозяев. Например, полупаразит Striga (колдунья трава) широко распространен в Африке, Южной Азии, Австралии на зерновых культурах, сорго, сахарном тростнике, образует на одном растении до 0,5 млн очень мелких семян. Семена паразитов весьма долговечны, что дает им возможность длительного «выжидания» контакта с растением-хозяином (семена Striga могут сохранять всхожесть в почве до 20 лет).
Громадная плодовитость, свойственная паразитам, получила название «закона большого числа яиц». Так, человеческая аскарида продуцирует в среднем 250 тыс. яиц за сутки, а за всю жизнь — свыше 60 млн. Подавляющее большинство яиц и зародышей паразитов гибнет, не выдержав воздействия различных факторов внешней среды, не попав в очередного хозяина, и только чрезвычайная плодовитость увеличивает шансы на выживание и завершение жизненного цикла хотя бы немногих потомков, поддерживая существование вида.
У ряда паразитов приспособления к умножению потомства проявляются и в виде партеногенеза, полиэмбрионии (клетки одного делящегося яйца дают начало множеству зародышей), бесполого размножения (почкование у пузырчатых стадий ленточных червей). Это приводит к чередованию поколений — полового и партеногенетического или полового и бесполых.
У многих паразитов чередование поколений сочетается со сменой двух или более хозяев, следовательно, одно поколение существует в одном хозяине, а другое — в другом. К примеру, ленточный червь, обитающий в кишечнике лисицы, откладывает там крошечные яички. Эти яички вместе с экскрементами лисицы попадают на землю. Как им теперь вернуться в кишечник какой-нибудь другой лисицы? Процесс возвращения включает в себя стадию промежуточного хозяина или несколько хозяев. Траву, на которую попали яйца ленточного червя, съедает заяц. В его организме яйца превращаются в личинки. Личинки вбуравливаются в ткани зайца и там переходят в покоящуюся стадию — цисты. В дальнейшем, если лисица поймает этого зайца, то цисты попадут в ее кишечник, там превратятся в молодого ленточного червя, и жизненный цикл начнется сначала. Многообразие и сложность жизненных циклов паразитов выработались как приспособление для передачи от одной особи хозяина к другой, для распространения.
Адаптация паразита к существованию на определенном хозяине включает также весьма тонкую биохимическую специализацию — выработку определенного набора ферментов, облегчающих проникновение в тело хозяина и использование поступающих от него веществ.
В ряде случаев паразиты сами становятся средой обитания других видов — возникает явление сверхпаразитизма, или гиперпаразитизма. Например, для паразита капустной белянки наездника (Apanteles glomeratus) известно более 20 видов вторичных паразитов из перепончатокрылых. Явления сверхпаразитизма в растительном мире сравнительно редки. К ним относятся случаи поселения одного вида омелы (Viscum meniliforme) на другом (V. orientale) в тропических лесах Индии.
В разных частях тела многоклеточного организма условия неоднородны. Для своих обитателей хозяин выступает как многообразная среда. Его паразиты специализируются к жизни в определенных органах и тканях. Они приурочены к определенному возрастному и физиологическому состоянию хозяина. Так, в волосах человека обитают одни разновидности вшей, тогда как в волосяном покрове других частей тела — совсем иные. В пищеварительном тракте кролика одновременно могут паразитировать несколько видов кокцидий, каждый из которых локализуется в определенных частях кишечника: Eimeria media — в начальном отделе тонкой кишки, Е. irresidua — в средней, а Е. magna — в последней ее петле, Е. piriformis — преимущественно в слепой кишке и т. д.
В Сибири, в лиственницах, тонкоусый еловый усач заселяет преимущественно прикомлевую часть до высоты примерно 1 м, лиственничная златка осваивает ствол выше, до 4—5 м, продолговатый короед распространяется по всей его средней части, а вершину и ветви заселяют короед-гравер и заболотник Моравица.
Большое число паразитов обитает не внутри, а на поверхности тела хозяина, выступающего в этом случае как часть внешней среды паразита, снабжая его пищей, предоставляя убежище, трансформируя микроклимат. Связь с хозяином эктопаразита может быть постоянной или временной. Одна из основных жизненно важных экологических задач для постоянных или длительно связанных эктопаразитов с хозяином — удержаться на теле хозяина. Типичные эктопаразиты в связи с этим обычно характеризуются наличием мощных органов прикрепления — присосок, крючьев, коготков и т. д., которые независимыми путями развиваются у самых разных по происхождению видов.
Живые организмы не только испытывают воздействия со стороны паразитов, но и энергично реагируют на них. Паразиты как обитатели живой среды должны преодолевать сопротивление организма хозяина, его защитные реакции. Это сопротивление паразитам носит название активного иммунитета. Здоровые, полноценные особи животных и растений обладают действенными защитными приспособлениями, которые не позволяют проникать в них патогенным организмам. Так, у животных защитной реакцией от вторжения посторонних организмов является выработка гуморального иммунитета, т. е. образование в крови хозяина специфических белковых веществ, антител, которые подавляют паразитов. Выработка иммунитета стимулируется токсинами паразита и часто предохраняет от повторных заражений. Устойчивость хвойных деревьев к нападению стволовых вредителей (жуков-короедов, усачей и др.) обеспечивается прежде всего выделением смолы, содержащей соединения, токсичные для этих насекомых. Ослабленные же деревья теряют сопротивляемость, подвергаются заселению насекомыми-вредителями, поражаются грибами и т. д.
В ряде случаев организм хозяина отвечает на вторжение паразита разрастанием окружающих его тканей, образованием своеобразной капсулы, которая изолирует паразита. Такие образования у растений называют галлами, а у животных — зооцецидия-ми. Нередко подобная изоляция приводит к гибели паразита. Чаще же, например, защитная роль зооцецидиев ограничивается локализацией причиняемого вреда в определенном участке тела хозяина, а сами паразиты используют их как дополнительные защитные образования. Особенно наглядно это выражено в возникновении галлов у растений (рис. 5.51).
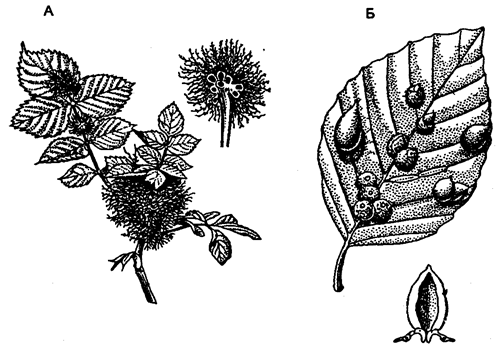
Рис. 5.51. Галлы на листьях (по Е. Страсбургу, 1962)
А — шиповник (Rosa conina); Б — бук (Fagus sytuatica)
Вызывающие их насекомые, клещи, нематоды и другие паразиты выделяют специфические вещества, которые стимулируют преобразование тканей или целых органов растений в галлы с камерой внутри, где их обитатель надежно защищен от высыхания, врагов и обеспечен пищей.
Как уже было отмечено, защитные реакции растения-хозяина состоят прежде всего в выработке иммунитета — невосприимчивости к заражению паразитами. Известны устойчивые к паразитным грибам сорта картофеля, зерновых и других сельскохозяйственных культур. К защитным приспособлениям относятся и особенности покровных тканей, которые затрудняют проникновение паразита (толстая кутикула, опушение и т. д.), особенности химического состава клеток и тканей. Например, доказано, что к грибным паразитам устойчивы растения, содержащие много эфирных масел, сапонинов, алкалоидов, многие галофиты с повышенным содержанием солей.
Отношения между паразитом и хозяином в растительном и животном мире на популяционном и видовом уровнях определенным образом уравновешены. Очевидно, паразит не может размножаться до такой степени, чтобы привести к вымиранию популяции хозяина и лишить себя «кормовой базы». Регулятором равновесия служит относительно медленное воздействие на хозяина некоторых паразитов, таких, как грибы, вызывающие ржавчину, головню, мучнистую росу на зерновых культурах, а иногда даже наблюдается некоторая биохимическая стимуляция роста хозяина со стороны паразита. Следовательно, паразиты, так же как и свободноживущие виды, имеют сложную систему приспособлений к своей среде обитания. Их строение и организация отражают специфику этой среды. Развиваются сходные типы приспособлений у представителей разных групп, ведущих паразитический образ жизни.
Дата добавления: 2016-02-24; просмотров: 1211;