Отношения организмов в биоценозах. Разнообразные формы биотических отношений, в которые вступают те или иные виды в биоценозе (конкуренция
Разнообразные формы биотических отношений, в которые вступают те или иные виды в биоценозе (конкуренция, комменсализм, мутуализм, хищник-жертва и др.), определяют основные условия их жизни в сообществе, возможности добывания пищи и завоевания нового пространства.
Прямые и косвенные межвидовые отношения по значению, которое они имеют для занятия видом в биоценозе определенного положения, по классификации В. Н. Беклемишева (1970), подразделяются на четыре типа: 1) трофические, 2) топические, 3) форические и 4) фабрические.
Трофические связи наблюдаются, когда один вид питается другим —либо их мертвыми остатками, либо продуктами их жизнедеятельности. Как стрекозы, ловящие на лету других насекомых, так и жуки-навозники, питающиеся пометом крупных копытных, и пчелы, собирающие нектар растений, вступают в прямую трофическую связь с видами, которые предоставляют им пищу. При конкуренции двух видов из-за объектов питания между ними возникает косвенная трофическая связь, вследствие того что деятельность одного отражается на снабжении кормом другого. Воздействие одного вида на поедаемость другого или доступность для него пищи расценивается так же, как косвенная трофическая связь между ними. Так, гусеницы бабочек-монашенок, объедая хвою сосен, облегчают короедам доступ к ослабленным деревьям.
Топические связи характеризуют любое физическое или химическое изменение условий обитания одного вида в результате жизнедеятельности другого. Данный вид связей отличается большим разноообразием. Топические связи заключаются в создании одним видом среды для другого (внутренний паразитизм или норовый комменсализм), в формировании субстрата, на котором поселяются или избегают поселяться представители других видов, во влиянии на движение воды, воздуха, изменение температуры, освещенности окружающего пространства, в насыщении среды продуктами насыщения и т. д. Морские желуди, поселяющиеся на коже китов, лишайники на стволах деревьев связаны прямой топической связью с организмами, представляющими им субстрат или среду обитания. Значительная роль в создании или изменении среды для других организмов принадлежит растениям. Из-за особенностей энергообмена растительность является мощным фактором перераспределения тепла у поверхности Земли и создания мезо- или микроклимата. Под пологом леса подлесок, напочвенный покров, животные находятся в условиях более выравненных температур, более высокой влажности воздуха и т. д. Хотя и в меньшей степени, травянистая растительность; также изменяет режим окружающего пространства. В результате положительных или отрицательных топических взаимоотношений одни виды определяют или исключают возможность существования в биоценозе других видов.
В биоценозе трофические и топические связи имеют наибольшее значение, составляют основу его существования. Эти типы| отношений удерживают друг возле друга организмы разных видов, объединяя их в сравнительно стабильные сообщества разных масштабов.
Форические связи — это участие одного вида в распространении другого. В роли транспортировщиков выступают животные. Как нами было отмечено ранее, перенос животными семян, спор, пыльцы растений называют зоохорией. Перенос же животными других, более мелких животных называют форезией (от лат. форас, — наружу, вон). Обычно перенос осуществляется с помощью специальных и разнообразных приспособлений. Форезия животных преимущественно распространена среди мелких членистоногих: например, у разнообразных групп клещей представляет собой один из способов пассивного их расселения. Она свойственна видам, для которых перенос из одного биотипа в другой жизненно необходим для сохранения или процветания. Так, многие летающие насекомые (рис. 11.9) — посетители скоплений быстро разлагающихся органических остатков (трупов, животных, куч гниющих растений и др.) — несут на себе гамазовых, уроподовых или тирогли-фоидных клещей, переселяющихся данным способом от одного скопления пищевых материалов к другому.
Рис. 11.9. Форезия клещей на насекомых (по W. Jacobs, М. Renner, 1974)
Жуки-навозники нередко ползают с поднятыми подкрыльями, так как не в состоянии их сложить из-за густо усеявших тело клещей. При помощи форезии на насекомых распространяются некоторые виды нематод. Ноги навозных мух иногда имеют вид ламповых щеток из-за обилия прикрепившихся особей рода Rhabditis. Форезия среди крупных животных практически не встречается.
Фабрические связи — это такой тип биоценотических отношений, в которые вступает вид, используя для своих сооружений (фабрикации) продукты выделения или мертвые остатки или даже живых особей другого вида (В. Н. Беклемишев, 1970). Например, птицы употребляют для постройки гнезд ветви деревьев, листья, траву, шерсть млекопитающих, пух и перья других видов птиц и т. д. Пчела-мегахила помещает яйца и запасы в стаканчики, которые сооружены из мягких листьев различных кустарников (акации, сирени, шиповника и др.).
Каждый конкретный вид из-за сложности межвидовых взаимоотношений может преуспевать не везде, где складываются подходящие для него условия физической среды. Отмечают физиологический и синэкологический оптимумы в распространении вида.
Физиологический оптимум — благоприятное для вида сочетание всех видов абиотических факторов, при котором возможны наиболее быстрые темпы роста и размножения. Синэкологический оптимум — биотическое окружение, при котором вид испытывает наименьшее давление со стороны врагов и конкурентов, что позволяет ему успешно размножаться. Физиологический и синзкологический оптимумы далеко не всегда совпадают. Большинство листостебельных растений — амфитолерантные формы с широким оптимумом от слабокислых до слабощелочных значений рН и с диапазоном толерантности от 3,5 до 8,5 рН при выращивании в одновидовых посевах. В естественном же распространении некоторые из них ограничены относительно низкими пределами рН. В таком случае их синэкологический оптимум не совпадает с физиологическим оптимумом (рис. 11.10).
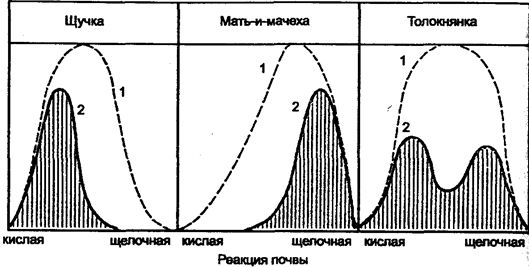
Рис. 11.10. Влияние рН на рост различных растений при выращивании
в одновидовых посевах и в условиях конкуренции (по В. Лархеру, 1978)
Примечание: кривые физиологического (1) и синэкологического (2) оптимумов
В отличие от видов, которые могут выдерживать конкуренцию в пределах своего физиологического оптимума, некоторые виды оттесняются в места с меньшей интенсивностью конкуренции, Тогда они полностью используют границы своей толерантности по отношению рН почвы. В результате, например, такой амфитолерантный вид растения, как толокнянка, распространен в большинстве своем на кислых и щелочных почвах.
Другой пример несовпадения физиологического и синэколо-гического оптимумов — массовое размножение вредителя зерновых культур гессенской мухи, после особенно суровых зим, которые должны были бы неблагоприятно сказаться на численности данного насекомого. В нормальные по условиям годы гессенскую муху сильно истребляют несколько видов ее естественных врагов — паразитических перепончатокрылых наездников. Из-за слабой морозоустойчивости в суровые зимы враги гессенской мухи погибают практически полностью. Это дает возможность вредителю восстановить собственную численность, уменьшенную низкими температурами в зимний период, и беспрепятственно размножаться в угрожающем для урожая количестве.
Межвидовые связи, формирующие биоценоз, обусловливают закономерные соотношения в нем видов, их экологических особенностей, численности, распределения в пространстве, или, можно сказать, позволяют создать определенную структуру биоценоза.
Экологические ниши
Экологической нишей называют положение вида, которое oi занимает в общей системе биоценоза, комплекс его биоценотических связей и требований к абиотическим факторам среды. Экологическая ниша отображает участие вида в биоценозе. При этом имеется в виду не территориальное его размещение, а функциональное проявление организма в сообществе. По выражению Ч. Элтона (1934), экологическая ниша — «это место в живом, окружении, отношение вида к пище и к врагам». Концепция экологической ниши оказалась весьма плодотворной для понимания законов совместной жизни видов. Помимо Ч. Элтона над ее развитием работали многие экологи, среди них Д. Гриннелл, Г. Хатчинсон, Ю. Одум и др.
Существование вида в сообществе определяется сочетанием и действием многих факторов, но в определении принадлежности организмов к той или иной нише исходят из характера питания этих организмов, из их способности добывать или поставлять пищу. Так, зеленое растение, принимая участие в сложении биоценоза, обеспечивает существование целому ряду экологических ниш. Это могут быть ниши, охватывающие организмы, питающиеся тканями корней или тканями листьев, цветками, плодами, выделениями корней и т. д. (рис. 11.11).
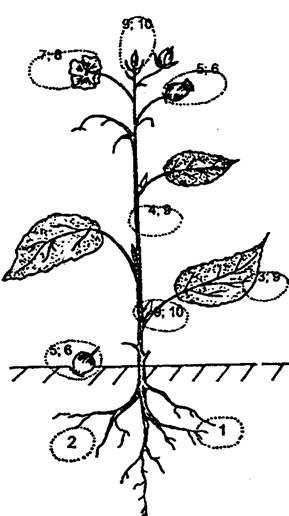
Рис. 11.11. Размещение экологических ниш, приуроченных к растению:
1 — корнееды; 2 — поедающие корневые выделения; 3 — листоеды; 4 — стволоеды, 5 — плодоеды; 6 — семяеды; 7 — цветкоеды; 8 — пыльцееды; 9 — сокоеды; 10 — почкоеды
(по И. Н. Пономаревой, 1975)
Каждая из этих ниш включает в себя разнородные по видовому составу группы организмов. Так, в экологическую группу корнеедов входят и нематоды, и личинки некоторых жуков (щелкуна, майского хруща), а в нишу сосущих соки растения — клопы, тли. Экологические ниши «стеблееды» или «стволоеды» охватывают большую группу животных, среди которых особенно многочисленны насекомые (древоточцы, древесинники, короеды, усачи и др.).
Следует отметить, что среди них имеются и такие, которые питаются только древесиной живых растений или только корой — те и другие принадлежат к разным экологическим нишам. Специализация видов в отношении пищевых ресурсов уменьшает конкуренцию, увеличивает стабильность структуры сообщества.
Существуют различные типы разделения ресурсов.
1. Специализация морфологии и поведения в соответствии с родом пищи: например, клюв у птиц может быть приспособлен для ловли насекомых, долбления отверстий, раскалывания орехов, разрывания мяса и др.
2. Вертикальное разделение, например, между обитателями полога и лесной подстилки.
3. Горизонтальное разделение, например, между обитателями различных микроместообитаний. Каждый из этих типов или их комбинация приводит к разделению организмов на группы, менее конкурирующих между собой, так как каждая из них занимает свою нишу. Например, существует разделение птиц на экологические группы, основанное на месте их питания: воздух, листва, ствол, почва. Дальнейшее подразделение этих групп в зависимости от основного типа пищи показано на рис. 11.12.
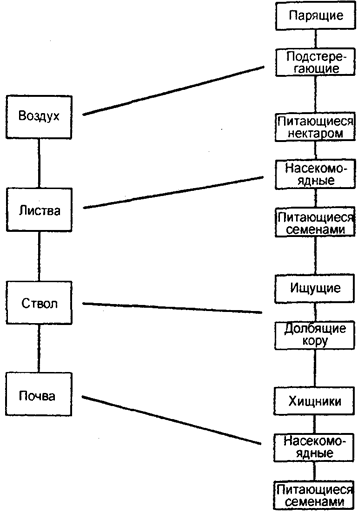
Рис. 11.12. Разделение птиц на экологические группы, основанное
на месте их питания: воздух, листва, ствол, земля
(по Н. Грину и др., 1993)
Специализация вида по питанию, использованию пространства, времени активности и другим условиям характеризуется как сужение его экологической ниши, а обратные процессы — как его расширение.
На сужение или расширение экологической ниши вида в сообществе большое влияние оказывают конкуренты. Сформулированное Г. Ф. Гаузе правило конкурентного исключения для близких по экологии видов может быть выражено таким образом, что два вида не уживаются в одной экологической нише. Выход из конкуренции достигается расхождением требований к среде, изменению образа жизни или, другими словами, является разграничением экологических ниш видов. В этом случае они приобретают способность сосуществовать в одном биоценозе. Так, в мангровых зарослях побережья Южной Флориды обитают самые разные цапли и нередко на одной и той же отмели кормятся рыбой до девяти разных видов. При этом они практически не мешают друг другу, так как в их поведении — в том, какие охотничьи участки они предпочитают и как ловят рыбу, — выработались приспособления, позволяющие им занимать различные ниши в пределах одной и той же отмели. Зеленая кваква пассивно поджидает рыбу, сидя на выступающих из воды корнях мангровых деревьев. Луизианская цапля делает резкие движения, взбалтывая воду и вспугивая затаившихся рыбок. Снежная цапля в поисках добычи медленно передвигается с места на место.
Наиболее утонченным способом лова пользуется красная цапля, которая сначала взбаламучивает воду, а затем широко раскрывает крылья, создавая тень. При этом, во-первых, она сама хорошо видит все происходящее в воде, а, во-вторых, вспугнутые рыбы принимают тень за укрытие, устремляются к нему, попадая прямо в клюв врага. Размеры большой голубой цапли позволяют ей охотиться в местах, не доступных для ее более мелких и коротконогих сородичей. Насекомоядные птицы в зимних лесах России, кормящиеся на деревьях, за счет разного характера поиска пищи также избегают конкуренции друг с другом. Поползни и пищухи собирают пищу на стволах. Поползни стремительно обследуют деревья, быстро схватывая насекомых, семена, оказавшиеся в крупных трещинах коры, а мелкие пищухи тщательно обшаривают на поверхности ствола малейшие щелки, в которые проникает их тонкий шиловидный клюв. В европейской части России существуют близкие виды синиц, изоляция которых друг от друга обусловлена различиями в местообитаниях, местах кормежки и размерах добычи. Экологические различия отражаются и в ряде небольших деталей внешнего строения, в том числе в изменениях длины и толщины клюва (рис. 11.13).
Зимой в смешанных стайках большие синицы ведут широкий поиск пищи на деревьях, в кустах, на пнях, а часто и на снегу. Синицы-гаички обследуют большей частью крупные ветви. Длиннохвостые синицы ищут корм на концах ветвей, а мелкие корольки тщательно обследуют верхние части крон хвойных.
Многочисленные отряды животных, питающихся травой, имеют в своем составе степные биоценозы. Среди них много крупных и мелких млекопитающих, таких, как копытные (лошади, овцы, козы, сайгаки) и грызуны (суслики, сурки, мышевидные). Все они составляют одну большую функциональную группу биоценоза (экосистемы) — травоядных животных. Вместе с тем исследования показывают, что роль этих животных в потреблении растительной массы не одинакова, так как они используют в своем питании разные составные части травяного покрова.
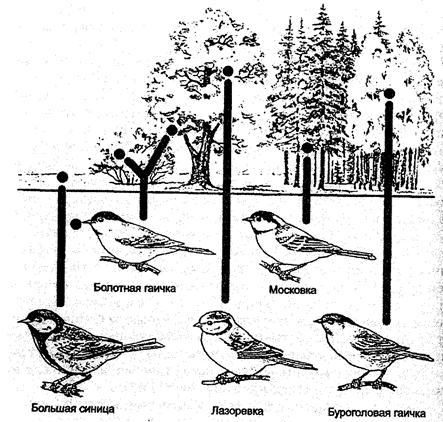
Рис. 11.13. Пищевые угодья у различных видов синиц
(по Е. А. Криксунову и др., 1995)
Так, крупные копытные (в настоящее время это домашние животные и сайгаки, а до освоения человеком степей — только дикие виды) лишь частично, выборочно выедают корм, главным образом высокие, наиболее питательные травы, откусывая их на значительной высоте (4—7 см) от поверхности почвы. Сурки, живущие здесь же, выбирают корм среди травостоя, изреженного и измененного копытными, поедая его, что было им недоступно. Сурки поселяются и кормятся только там, где нет высококотравья. Более мелкие животные — суслики — предпочитают собирать корм там, где еще сильнее нарушен травостой. Здесь они собирают то, что осталось от кормления копытных и сурков. Между этими тремя группами травоядных животных, образующих зооценоз, наблюдается разделение функций в использовании биомассы травянистого покрова. Отношения, сложившиеся между этими группами животных, не носят конкурентного характера. Все эти виды животных используют разные составные части растительного покрова, «доедая» то, что не является доступным другим травоядным. Разнокачественное участие в поедании травостоя или размещение организмов по разным экологическим нишам обеспечивает более сложную структуру биоценоза на данной территории, обеспечивая более полное использование условий жизни в природных экосистемах и максимальное потребление ее продукции. Совместное существование этих животных характеризуется не только отсутствием конкурентных связей, а наоборот, обеспечивает высокую их численность. Так, отмечающееся в последние десятилетия увеличение сусликов и их расселение — результат усиления выпаса домашних животных в степных районах в связи с увеличением поголовья скота. В местах же, лишенных выпаса (например, заповедные земли), наблюдается сокращение численности сурков и сусликов. На участках с быстрым отрастанием трав (особенно на высокотравных участках) сурки уходят совсем, а суслики остаются в незначительном количестве.
Экологические ниши видов изменчивы в пространстве и во времени. Нередко в биоценозе один и тот же вид в разные периоды развития может занимать различные экологические ниши. Так, головастик питается растительной пищей, а взрослая лягушка—типичное плотоядное животное, и им свойственны различные экологические ниши и специфические трофические уровни. Разными экологическими нишами зимой и летом в связи с миграциями характеризуются и перелетные птицы. У насекомоядных птиц зимние экологические ниши отличаются от летних. В разные экологические ниши входят личинки оводов, паразитирующие на крупных млекопитающих, и их взрослые особи, не принимающие совсем пищу, или некоторые бабочки, у которых чрезвычайно активными являются гусеницы, пожирающие листья, хвою, а взрослые потребляют нектар или вообще не питаются. То же и у майского хруща: взрослое насекомое относится к экологической нише листоедов, а личинка — корнеед. Среди водорослей имеются виды, которые функционируют то как автотрофы, то как гетеротрофы, тем самым занимая в определенные периоды жизни те или иные экологические ниши.
У растений, живущих в одном ярусе, экологические ниши сходны, что способствует ослаблению конкуренции между растениями разных ярусов и обусловливает освоение ими различных экологических ниш. В биоценозе разные виды растений занимают разные экологические ниши, что ослабляет межвидовую конкурентную напряженность. Один и тот же вид растений в различных природных зонах может занимать разные экологические ниши. Так, сосна и черника в бору-черничнике, водные растения (рдесты, кубышка, водокрас, ряски) поселяются вместе, но распределяются по различным нишам. Седмичник и черника в лесах умеренной полосы являются типичными теневыми формами, а в лесотундре и тундре растут на открытых пространствах и становятся световыми. На экологическую нишу вида оказывают влияние межвидовая и внутривидовая конкуренции.
При наличии конкуренции с близкородственными или экологически сходными видами зона местообитания сокращается до отщ мальных границ (рис. 11.14), т. е. вид распространяется в наиболе< благоприятных для него зонах, где он обладает преимуществом пс сравнению со своими конкурентами. Если межвидовая конкуренция сужает экологическую нишу вида, не давая проявиться в полном объеме, то внутривидовая конкуренция, напротив, способствует расширению экологических ниш. При возросшей численностщ вида начинается использование дополнительных кормов, освоение новых местообитаний, появление новых биоценотических связей.
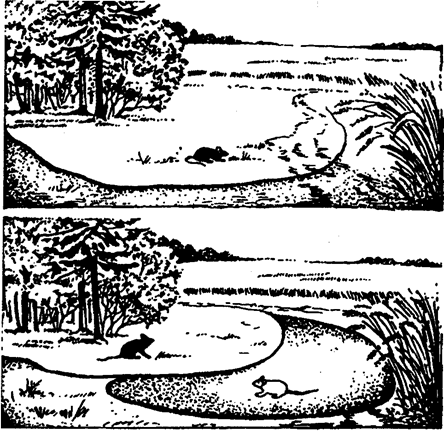
Рис. 11.14. Разделение мест обитания вследствие конкуренции
(по Е. А. Криксунову, 1995)
Дата добавления: 2015-12-01; просмотров: 1380;