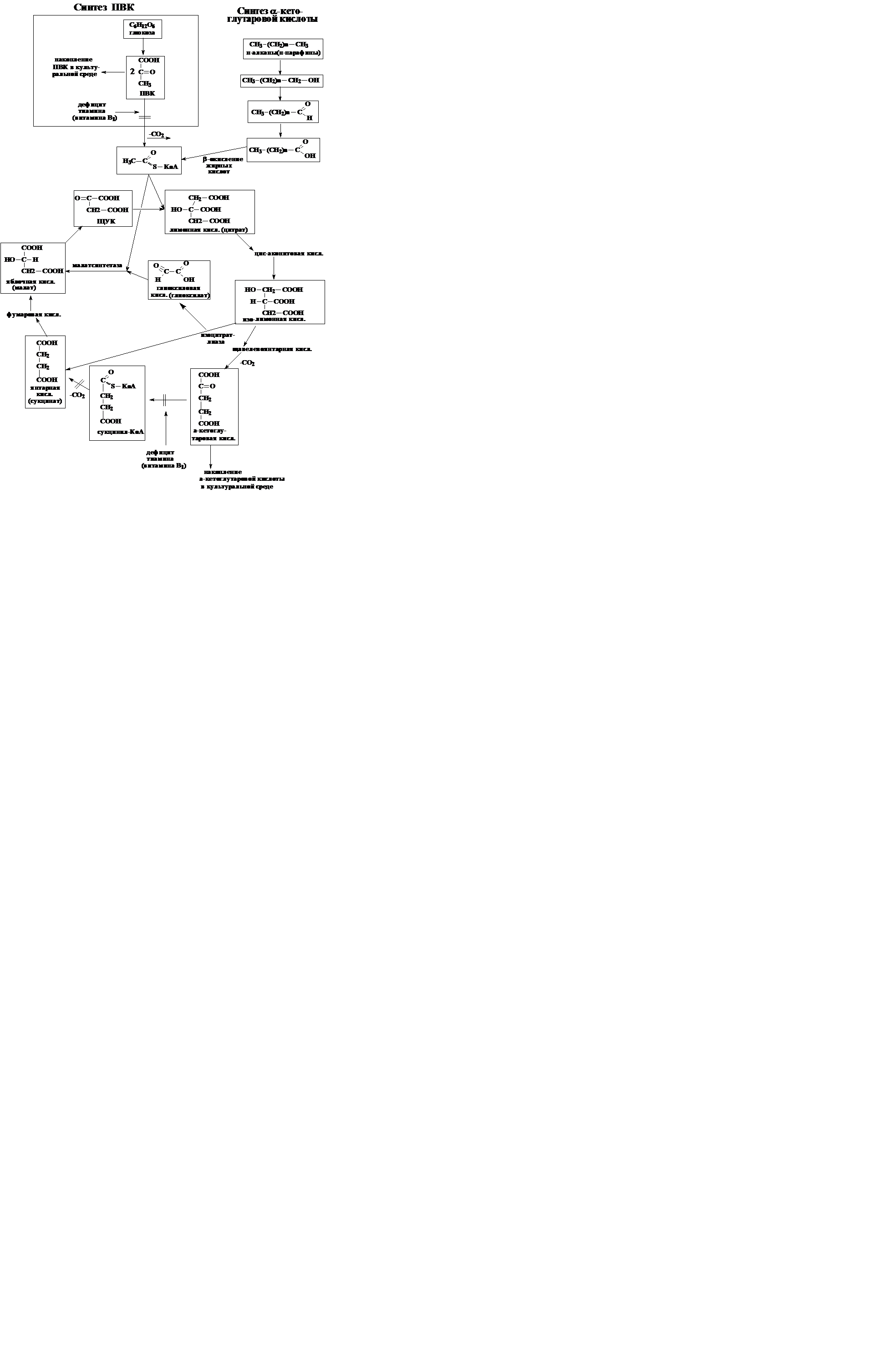
Получение кетокислот (пировиноградной и кетоглутаровой).
Как известно тиамин (витамин В1) в форме пирофосфатного производного является простетической группой ферментов, осуществляющих реакции расщепления и синтеза углерод-углеродных связей, находящихся между карбоксильной и карбонильной группами в молекулах α-кетокислот. Примерами таких процессов являются реакции окислительного декарбоксилирования ПВК и α-кетоглутаровой кислоты в цикле Кребса.
Большинство природных микроорганизмов способны самостоятельно синтезировать всю молекулу тиамина, однако методом мутагенеза и селекции получены виды неспособные к этому – тиамингетеротрофыили более правильно тиаминауксотрофы,нуждающиеся в добавлении тиамина к питательным средам.
При этом в зависимости от субстрата, на котором проводится культивирование, дефицит тиамина по разному отражается на обмене веществ у тиамингетеротрофов, поскольку может происходить избирательное блокирование тех или иных процессов карбоксилирования и декарбоксилирования.
Так, при окислении глюкозы тиамингетеротрофными дрожжами рода Candida промежуточным продуктом ее превращения в ацетил-КоА является пировиноградная кислота, окислительное декарбоксилирование которой осуществляется полиферментным комплексом, включающим ТПФ-зависимую
пируватдекарбоксилазу.
Дрожжи, выращиваемые на глюкозной среде с дефицитом по тиамину, не могут нормально осуществлять реакцию декарбоксилирования ПВК, что приводит к торможению или блокированию образования ацетил-КоА. Следствием дефицита тиамина является выделение ПВК в культуральную среду, нарушение функционирования цикла Кребса, снижение биосинтетических процессов в клетке, приводящее к прекращению роста культуры.
При росте на среде с н-алканами ацетил - КоА образуется в результате
β-окисления алифатических жирных кислот, образующихся из н-алканов, минуя стадию образования ПВК. Если в культуральной среде создается лимит по тиамину, то процессы окисления в цикле Кребса прекращаются на стадии образования α-кетоглутаровой кислоты. Это связано с тем, что блокируется функционирование другого тиаминзависимого ферментного комплекса - α-кетоглутаратдегидрогеназы, катализирующей окислительное декарбоксилиро-вание α-кетоглутаровой кислоты и образование сукцинил-КоА. Следствием дефицита тиамина является накопление α-кетоглутаровой кислоты в клетке, ее выделение в среду и замедление роста клеток. Однако зависимость клеток от тиамина в данном случае гораздо меньше. Даже при глубоком дефиците тиамина в среде - 3-4 ·10-4 мкг/мл, что соответствует его содержанию в водопроводной воде - дрожжи растут и выделяют в среду α-кетоглутаровую кислоту до полного потребления н-алканов. Это объясняется тем, что в клетках часть молекул ацетил-КоА метаболизируется по глиоксилатному циклу. Глиоксилатный цикл позволяет клетке не только получать энергию (АТФ), и самое главное, обеспечивает расширенный ресинтез (2 молекулы на цикл) щавелевоуксусной кислоты, восполняя ее утечку с α-кетоглутаровой кислотой из ЦТК.
α-Кетоглутаровая кислота - ценный химический реагент, используемый для производства глутаминовой кислоты и некоторых гетероциклических соединений. Промышленное производство α-кетоглутаровой кислоты микробиологическим путем налажено во многих странах.
ЛЕКЦИЯ №4
Дата добавления: 2016-01-30; просмотров: 2457;