ЭВОЛЮЦИОННЫЕ ПРЕОБРАЗОВАНИЯ МОЗГА
Строение мозга у животных разных видов разнообразно. И хотя, как следует из рис. 19.3, родственные виды, например среди ракообразных или млекопитающих, имеют общие черты в строении мозга, между собой они мало похожи. Именно с появлением нервной системы организмы получили возможность быстрой адаптации к быстро меняющимся факторам среды, что, безусловно, давало преимущества в борьбе за существование. У предков млекопитающих, как и у современных рептилий, кора больших полушарий была очень слабо дифференцирована [Батуев, 1979]. Но на пути от рептилиеподобных предков млекопитающих до ныне живущих млекопитающих произошло значительное увеличение коры мозга по сравнению с другими структурами мозга (рис. 19.4), изменение количества (см. рис. 19.3 В), размеров и формы корковых областей, их модульного строения и связей [Kaas, 1987], произошло увеличение переднего мозга и, как предполагается [McLean, 1993; Povinelli, Preuss, 1995], перемещение главенствующей роли от стриарного комплекса сначала к лимбической системе, а затем к неокортексу и особенно к префронтальной области коры (см. рис. 19.3 Б).
Важной чертой эволюции млекопитающих является уже упомянутое увеличение неокортекса, при этом степень этого увеличения отличает приматов от остальных млекопитающих, а человека – от остальных приматов. В частности, соотношение площади неокортекса у мыши, макаки и человека составляет 1 : 100 : 1000 соответственно [цит. по: Rakic, 1995], а соотношение объёмов коры мозга и спинного мозга у крыс и у человека – 31 : 35 и 77 : 2, соответственно (см. рис. 19.4) [Swanson, 1995]. Такая экспансия неокортекса, как полагают, связана с модификацией регуляторных генов, определяющих митотическое деление клеток вентрикулярного (пролиферативного) слоя коры [Rakic, 1995]. При этом, по-видимому, кора развивалась под влиянием внутрикорковых или непрямых таламических подкорковых влияний [Krubitzer, 1995]. Такое развитие коры привело к возникновению новых мощных приспособительных функций [Innocenti, Kaas, 1995]. Можно предположить, что именно это явилось основой такого свойства нервной системы, которое У. Матурана обозначил как способность нервной системы «взаимодействовать со своими собственными внутренними состояниями, как если бы те были независимыми сущностями» [Матурана, 1995, с. 104]. В таком «взаимодействии» нервной системы со своими собственными состояниями может лежать ключ и к пониманию онтологической сути субъективных состояний – «переживаний», «мыслей», «образов», – а также языка и мышления.
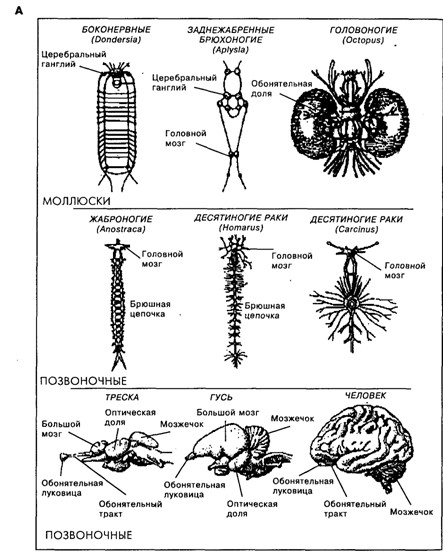
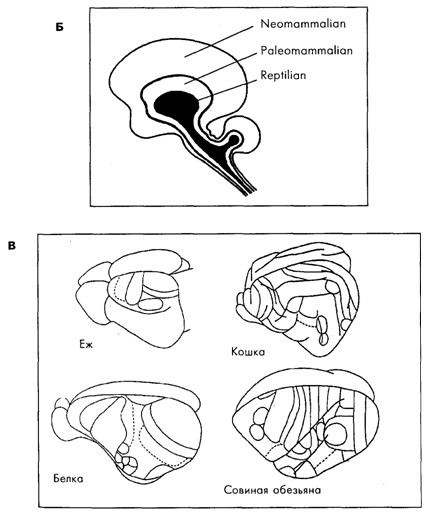
Рис. 19.3. А.Схематичное представление мозга животных, относящихся к линиям моллюсков, членистоногих и хордовых (масштабы разные)[по Кэндел, 1980]
Б. Условное представление триединой эволюции трёх основных мозговых ансамблей, приведших к образованияю переднего мозга у людей и у других развитых млекопитающих [McLean, 1993].
В . Корковые поля у примитивных млекопитающих (ёж), у млекопитающих с незначительно более развитой нервной системой (белка) и у двух млекопитающих с умеренно развитым мозгом (кошка и совиная обезьяна) [Kaas, 1987]
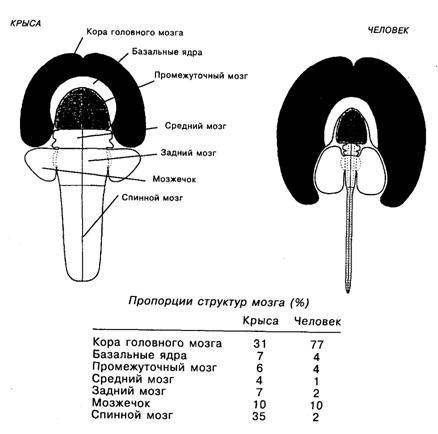
Рис. 19.4. Сравнение размеров разных отделов цетральной нервной системы крысы и человека. Пропорции между областями мозга соблюдены [Swonson, 1995]
Внутрикорковые процессы рассматриваются рядом авторов в качестве нейронной основы сознания [Маунткасл, 1981; Эделмен, 1981; Eccles, 1992]. Так, например, в концепциях В. Маунткасла и Дж. Эделмена сознание возникает на основе повторного входа «сигнализации» (см. гл. 11), при этом, согласно одним взглядам, данный процесс происходит в модульных распределённых сетях [Маунткасл, 1981], а согласно другим – в вырожденной группе нейронов [Эделмен, 1981]. По мнению же Дж. Экклса, сознание – побочный эффект, возникший в эволюции неокортекса, наблюдающийся только у млекопитающих и сводящийся к взаимодействию мозга и ментального мира в множестве пресинаптических окончаний корковых нейронов, которые образуют морфологическую единицу – дендрон [Eccles, 1992].
Дата добавления: 2016-01-29; просмотров: 836;