Гены иммуноглобулинов
Прежде всего мы должны рассмотреть, как организованы гены ИГ у будущих лимфоцитов до того, как эти клетки стали отличаться друг от друга, т. е. до того, как образовались разные их клоны. Очевидно, что так же организованы гены ИГ и у всех других клеток организма, у тех, которые дифференцируются в иных направлениях и никогда не станут лимфоцитами.
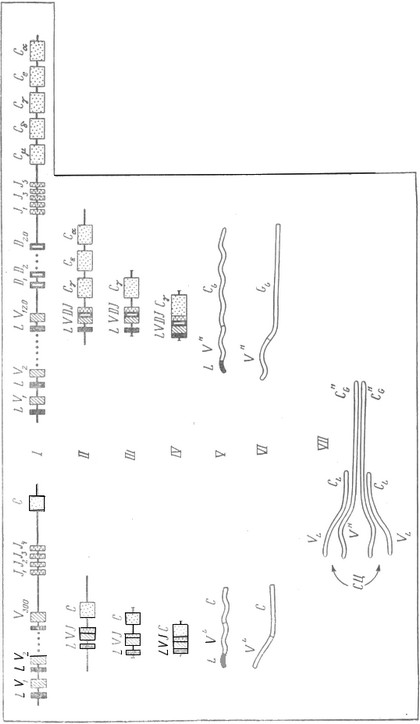
Гены легких цепей расположены в двух разных хромосомах, а гены тяжелых – в третьей. Каждая из них в диплоидной клетке, естественно, представлена парой гомологичных хромосом. Это в принципе создает возможность для образования в одной клетке не одного, а двух видов легких цепей (каппа‑ и лямбда‑типа), а с учетом возможных различий гомологичных хромосом и пары хромосом с генами тяжелых цепей – восьми видов ИГ. Фактически же в одном лимфоците всегда образуется только один вид ИГ. Это достигается путем так называемого аллельного исключения. Если в ИГ‑продуцирующих клетках перестраивается и затем активируется ген легкой цепи каппа в первой хромосоме, то гены легких цепей каппа в гомологичной хромосоме не перестраиваются и не активируются, а как бы исключаются из состава генов данной клетки – родоначальницы клопа. Такое же исключение происходит и во всех потомках этой клетки, т. е. во всем клопе. Если в данном лимфоците активируется ген каппа‑цепи в одной из хромосом, то не включаются и оба гена лямбда‑цепей в другой паре хромосом. Наоборот, включение одного из лямбда‑генов выключает тем самым оба каппа‑гена. В итоге из четырех возможных активным в лимфоците остается только один ген легких цепей. Аналогичным образом активируется ген тяжелой цепи только в одной из двух гомологичных хромосом. В другой гомологичной хромосоме аллельный ген тяжелой цепи исключается. Благодаря аллельному исключению достигается то, что в клетках каждого клона работают только два гена ИГ – одной легкой цепи (каппа или лямбда) и одной тяжелой – и каждый лимфоцит продуцирует только один вид антител. Механизм аллельного исключения неизвестен. Он представляет в генетике один из очень немногих примеров такого рода,
«Ген» каждой цепи ИГ представляет собой сложную многокомпонентную систему, располагающуюся вдоль хромосомы на значительное расстояние, в десятки тысяч пар оснований (килобаз). Называть эту систему одним геном было бы так же неверно, как и считать ее десятком или сотней генов. Это именно система, состоящая из множества вариабельных и нескольких константных последовательностей, которые собираются в один ген иммуноглобулина только, в результате довольно сложных преобразований, которые происходят при появлении первых лимфоцитов, еще в эмбриогенезе.
Система гена легких цепей каппа состоит из следующих компонентов (рассматривая их в порядке считывания слева направо). В начале системы (слева) находится большая группа участков ДНК, кодирующих вариабельную часть гена. Каждый из них содержит 291 пару нуклеотидов и кодирует соответственно 97 аминокислот. Всего таких участков около трехсот. Далее (правее) через значительное расстояние (около 2700 пар нуклеотидов) находится группа небольших участков «джей» (их четыре или пять), кодирующих по 13 аминокислот, соединяющих вариабельную и константную части ИГ. Наконец, дальше (еще правее) располагается участок, кодирующий константную часть ИГ.
Во время дифференцировки лимфоцитов в одной из хромосом, несущих систему каппа‑гена легких цепей, происходит сближение одного из вариабельных участков (V ) с одним из участков «джей» (J ) и с участком, кодирующим константную часть гена (С ). Пока не до конца ясно, каков конкретный механизм сближения: это может быть делеция, т. е. выбрасывание части ДНК так, что одна из F‑последовательностей оказывается рядом с одним из J . Ho может быть это и транспозиция, т. е. перенос одной из V к одному из J. Важно то, что процесс этот случаен и рядом с любой из V (их 300) может оказаться любая из J (их 4). В итоге создаются комбинации, общее возможное число которых нетрудно подсчитать: 300 × 4 = 1200. Это число и означает количество теоретически возможных вариантов каппа‑гена легких цепей. В лямбда‑генах такой принципиальной перестройки не происходит, но самих этих генов у мыши 20 и если в лимфоците образуется легкая лямбда‑цепь, то одна из двадцати возможных.
Приблизительно так же, как и каппа‑ген, устроена и система гена тяжелых цепей. Различия состоят в том, что число вариабельных участков там меньше‑(около 120), но зато между ними и «джей» находится еще 20 вариантов элемента D . Кроме того, в систему гена тяжелой цепи входит не один, а несколько константных участков, от которых, однако, антиген‑узнающая специфичность антител не зависит. При случайном сближении элементов V , J и D (путем делеции ДНК между ними или путем их транспозиции) теоретически возможно следующее количество вариантов: 120V × 20D × 4J = 9600.
В итоге в лимфоците случайным образом создаются два гена – для легких и для тяжелых цепей. Так как в молекуле ИГ (антитела) свойства центра связывания антигена определяются вариабельными частями и легкой и тяжелой цепей совместно, то в принципе число вариантов строения такого центра равно числу возможных сочетаний и превышает десять миллионов: 1200 × 9600 = 11,5 · 108. Действительно ли число возможных вариантов так велико, как это представляется из этих расчетов, точно пока неизвестно. Полагают, что оно несколько меньше.
Механизм делений, или транспозиций, также неизвестен, но подобный процесс определенно происходит, так как еще в первых работах Тонегавы было показано, что строение гена ИГ в лимфоците отличается от строения этого участка в ДНК недифференцированной или иначе дифференцированной клетки. Пока это единственный известный случай, когда в ходе дифференцировки происходит непременная, хотя и случайная перестройка генома. Что же касается деталей и точных механизмов такой перестройки, то на них сейчас обращено внимание стольких квалифицированных исследователей, что решение проблемы – это вопрос только времени.
Выше мы говорили, что система гена тяжелых цепей содержит не один, а несколько константных частей. Действительно, по ходу дифферепцировки лимфоцита в нем происходит смена этих частей. Сначала в состав гена входит только ближайшая константная часть и синтезируется антитело класса М, которое располагается на поверхности лимфоцита и является как бы выставкой, образцом его продукции. Если антиген окажется комплементарным этому образцу, то их соединение стимулирует лимфоцит к размножению и синтезу антител следующего класса (используется следующая константная часть класса G). Эти антитела уже секретируются в кровь и создают иммунитет.
Так, в результате преобразований в относительно небольшой части генома создается более миллиона различных клонов лимфоцитов, способных создавать иммунитет практически против любого антигена, случайно или искусственно попавшего в организм. Количество этих клонов намного превышает общее число генов. Оно, конечно, никак не могло быть получено «обычным путем», т. е. за счет наличия в геноме многих различных генов иммуноглобулинов и включения одного из них. Перестройка генов для образования разных антител – еще один пример того, что эволюция способна создавать такие «чудеса», которые не может предугадать ничья фантазия. Биологический смысл появления в эволюции подобного механизма очевиден – создание большого и случайного разнообразия за счет относительно небольшого участка ДНК.
Можно ли ожидать, что механизм, подобный этому, встретится и в других дифференцировках? Подобный механизм может оказаться целесообразным только там, где существенна не определенность, а разнообразие, даже случайное. Может быть, мы встретимся с чем‑то подобным при изучении связей между отдельными нервными клетками мозга. А может быть, нечто похожее происходит при образовании пятнистой окраски, там, где положение пятен должно быть случайным. А может быть, перестройка генетической системы синтеза ИГ – это уникальный механизм и мы не встретим его больше нигде.
Дата добавления: 2016-01-26; просмотров: 914;