ВТОРИЧНОЕ СТРОЕНИЕ КОРНЯ
У голосеменных и двудольных растений первичное строение сохраняется недолго и выше зоны всасывания сменяется вторичным. Вторичное утолщение корня происходит за счет деятельности вторичных боковых меристем – камбия и феллогена.
Камбий возникает в корнях из меристематических прокамбиальных клеток в виде прослойки между первичными ксилемой и флоэмой. На поперечном срезе камбий имеет вид однослойных, вогнутых внутрь дуг. Когда дуги камбия достигают перицикла, его клетки тоже начинают делиться, образуя клетки, соединяющие дуги камбия в единое камбиальное кольцо, окружающее первичную ксилему. В зависимости от числа флоэмных тяжей одновременно закладываются две или более зоны камбиальной активности.
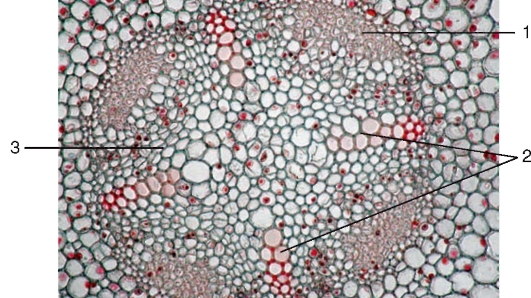
Рис.Переход от первичного к вторичному строению корня: 1 - участки флоэмы; 2 - четыре луча ксилемы; 3 - дуги камбия
Камбий откладывает внутрь слои вторичной ксилемы (древесины) и наружу вторичную флоэму (луб). Если этот процесс длится долго, то корни достигают значительной толщины.
Между элементами вторичной ксилемы и внутренним слоем вторичной флоэмы находится пучковый камбий; части перициклического происхождения, огибающие первичные лучи ксилемы, называют камбием межпучковьм. Межпучковый камбий образует паренхимные клетки, отходящие от радиальных лучей первичной ксилемы, называемых сердцевинными (паренхимными) лучами. Паренхимные лучи, образованные межпучковым камбием, изначально представляют собой первичные лучи. Вторичной ксилемы всегда бывает больше, чем вторичной флоэмы, и она оттесняет камбий наружу. При этом дуги камбия сначала выпрямляются, а затем принимают выпуклую форму. В результате деятельности камбия первичная флоэма оттесняется наружу и сдавливается. Звезда первичной ксилемы остается в центре корня, ее лучи могут сохраняться в течение длительного времени, но чаще центр корня заполняется вторичной ксилемой, и первичная ксилема становится незаметной. Сердцевинные лучи обеспечивают связь между ксилемой и флоэмой корня, по ним происходит радиальный транспорт различных соединений.

Рис. Поперечный срез корня тыквы (вторичное строение): 1 – первичная ксилема; 2 – вторичная ксилема; 3 – камбий; 4 – вторичная флоэма; 5 – первичный сердцевинный луч; 6 – пробка; 7 – паренхима вторичной коры.
Ткани первичной коры не могут следовать за вторичным утолщением и обречены на гибель. Они заменяются вторичной покровной тканью – перидермой, которая может растягиваться на поверхности утолщающегося корня благодаря работе феллогена. Феллогензакладывается в перицикле и начинает откладывать наружу пробку, а внутрь - феллодерму. Первичная кора, отрезанная пробкой от внутренних живых тканей центрального осевого цилиндра, отмирает и сбрасывается.
Клетки феллодермы и паренхима, образовавшаяся за счет деления клеток перицикла, образуют паренхиму вторичной коры, окружающую проводящие ткани. Снаружи корни вторичного строения покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев. Все ткани, расположенные от перидермы до камбия, входят в понятие «вторичная кора».
Многолетние корни древесных растений в результате длительной активности камбия нередко сильно утолщаются. Вторичная ксилема у таких корней сливается в сплошной цилиндр, окруженный снаружи кольцом камбия и сплошным кольцом вторичной флоэмы.
МЕТАМОРФОЗЫ КОРНЕЙ
Если корни выполняют особые функции, их строение меняется. Резкое, наследственно закрепленное видоизменение органа, вызванное сменой функций, носит название метаморфоза. Видоизменения корней очень разнообразны.
Микориза.Это симбиотическое взаимодействие гиф гриба и корневых окончаний растения. Микориза образуется на сосущих корнях в зоне поглощения. Грибы, живущие на корнях растений, используют органические вещества, синтезируемые зеленым растением, и поставляют растению из почвы воду и минеральные вещества. Благодаря сильно разветвленным гифам гриба у растения увеличивается всасывающая поверхность. Однако иногда симбиоз может переходить в паразитизм, например, когда гифы гриба наносят растению вред, угнетая его рост, или когда растение начинает использовать клеточное содержимое гиф гриба, проникших в коровую паренхиму растения.
Различают два основных типа микоризы. Гифы эктотрофной микоризы образуют чехол, окутывающий корень снаружи. Эктомикориза широко распространена у деревьев и кустарников. Эндотрофная микориза встречается в основном у травянистых растений. Эндомикориза находится внутри корня, гифы внедряются в клетки коровой паренхимы. Микотрофное питание очень широко распространено. Некоторые растения, например орхидные, вообще не могут существовать без симбиоза с грибами.
Клубеньки.Наличие клубеньков характерно для представителей семейства бобовых (люпина, клевера и др.). Клубеньки образуются в результате проникновения через корневые волоски в кору корня бактерий рода Rhizobium. Бактерии вызывают усиленное деление паренхимы, которая образует выросты бактероидной ткани на корне - клубеньки. Кроме того, они фиксируют атмосферный молекулярный азот и переводят его в связанное состояние в виде азотистых соединений, усваиваемых растением. Бактерии, в свою очередь, используют вещества, находящиеся в корнях растения. Такой симбиоз очень важен для почв и используется в сельском хозяйстве для их обогащения азотистыми веществами.
На корнях бобовых и других растений возникают особые образования - клубеньки, в которых поселяются бактерии. Эти бактерии способны фиксировать атмосферный молекулярный азот, переводя его в связанное состояние. Часть азотистых соединений, образованных таким путем, усваивает высшее растение-хозяин, отдавая при этом бактериям углеводы.
Воздушные корниобразуются у многих тропических эпифитных однодольных из семейств орхидных, ароидных, бромелиевых.Эпифиты поселяются на других растениях, но не паразитируют на них, а используют как подпорку для поднятия вверх, к свету. Воздушные корни свободно висят в воздухе и приспособлены к поглощению атмосферной влаги. На их поверхности образуется веламен. Веламен, как и ризодерма, образуется из протодермы, но это многослойная ткань. Клетки веламена отмирают, их стенки имеют сетчатые или спиральные утолщения. Через поры и сквозные отверстия в клеточных стенках влага проникает капиллярным путем. Изнутри веламен подстилается экзодермой со сложно построенными пропускными клетками, через которые вода передается в клетки коры и осевого цилиндра. Веламен встречается и у некоторых наземных однодольных (кливия, хлорофитум), в этом случае он выполняет функцию механической защиты и предохраняет растение от потери воды из первичной коры.
Дыхательные корни, илипневматофоры образуются у некоторых тропических древесных растений, живущих в условиях недостатка кислорода (таксодиум, или болотный кипарис; растения мангровых зарослей, обитающие по болотистым берегам океанических побережий). Пневматофоры растут вертикально вверх и высовываются над поверхностью почвы. Через систему отверстий в этих корнях, связанных с аэренхимой, воздух поступает в подводные органы.
У некоторых растений для поддержания побегов в воздушной среде образуются дополнительные опорные корни. Они отходят от горизонтальных ветвей кроны и, достигнув поверхности почвы, интенсивно ветвятся, превращаясь в столбовидные образования, поддерживающие крону дерева (столбовидные корни баньяна).
Ходульные корни отходят от нижних участков стебля, придавая стеблю устойчивость. Они образуются у растений мангровых зарослей, растительных сообществ, развивающихся на затопляемых во время прилива тропических берегах океанов, а также у кукурузы. У фикуса каучуконосного образуются досковидные корни. В отличие от столбовидных и ходульных, они являются по происхождению не придаточными, а боковыми корнями.
Побеги плюща, стремясь к солнцу, обвивают другие растения или прикрепляются к стенам при помощи корней - прицепок. У паразитических растений корни видоизменяются в присоски – гаустории, которые внедряются в ткани других растений и поглощают из клеток воду и питательные вещества
У многих луковичных и корневищных растений образуются втягивающие, или контрактильные корни. Они могут укорачиваться и втягивать побег в почву на оптимальную глубину на время летней засухи или зимних морозов. Втягивающие корни имеют утолщенные основания с поперечной морщинистостью.
Корневые клубни.У многих двудольных и однодольных растений в результате метаморфозы придаточных корней образуются корнеклубни (чистяка весеннего, любки двулистной, ятрышника пятнистого и т.д.).
Корневые клубни имеют ограниченный рост и приобретают овальную или веретеновидную форму. Такие клубни выполняют запасающую функцию, а поглощение почвенных растворов за них выполняют хорошо ветвящиеся всасывающие корни. У некоторых растений, таких как георгин, лилейник, корнеклубни выполняют запасающую функцию только в определенной части (базальной, срединной) а остальная часть клубня имеет типичное строение корня.
Такие корнеклубни могут выполнять как запасающую, так и всасывающую функции.
Корнеплоды.В образовании корнеплода могут участвовать различные части растения, например, разросшаяся базальная часть главного корня, утолщенный гипокотиль и др. Короткокорнеплодные сорта представителей семейства капустных (редис, репа) имеют плоский или округлый клубень, большая часть которого представлена разросшимся гипокотилем. Такие корнеплоды имеют вторичное анатомическое строение при диархной первичной ксилеме и хорошо развитой вторичной ксилеме, выполняющей запасающую функцию (рис. 3.15, 3.16). Клубень длиннокорнеплодных сортов представителей семейства сельдерейных
Рис. 3.19.Корнеплоды моркови (а, б),репы (в, г),свеклы (д, е, ж).На поперечных срезах (б, г, е)ксилема показана красным. Пунктирной линией обозначена граница стебля и корня
Дата добавления: 2016-01-03; просмотров: 13197;